

The Effect of Wnt Pathway Modulators on Human iPSC-Derived Pancreatic Beta Cell Maturation
- PMID: 31139151
- PMCID: PMC6518024
- DOI: 10.3389/fendo.2019.00293
The Effect of Wnt Pathway Modulators on Human iPSC-Derived Pancreatic Beta Cell Maturation
Abstract
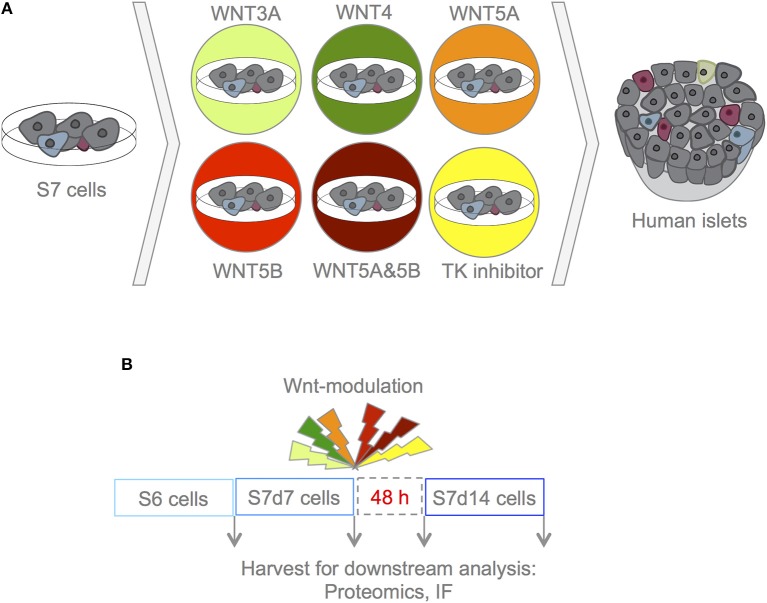
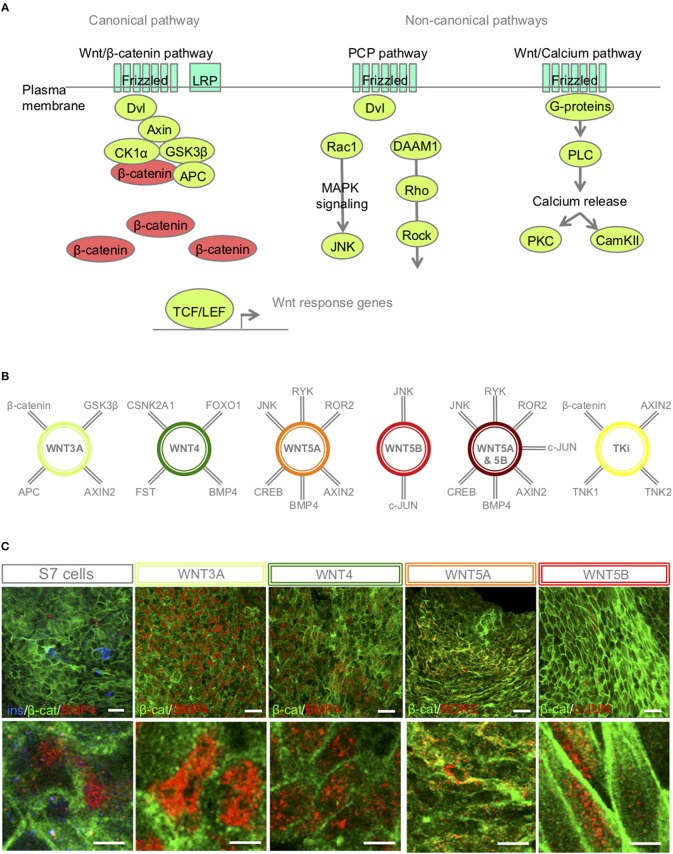
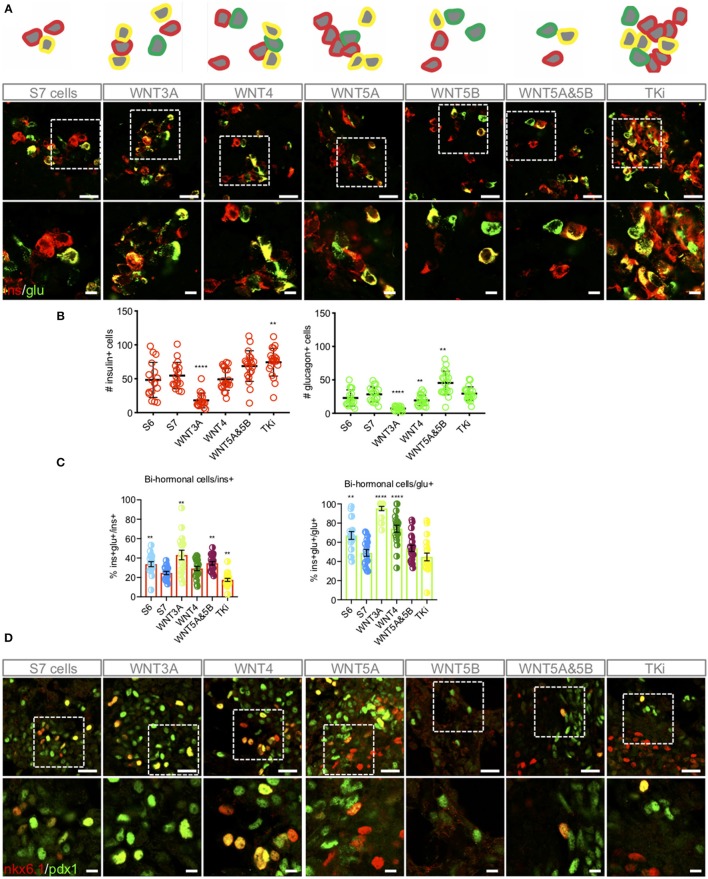
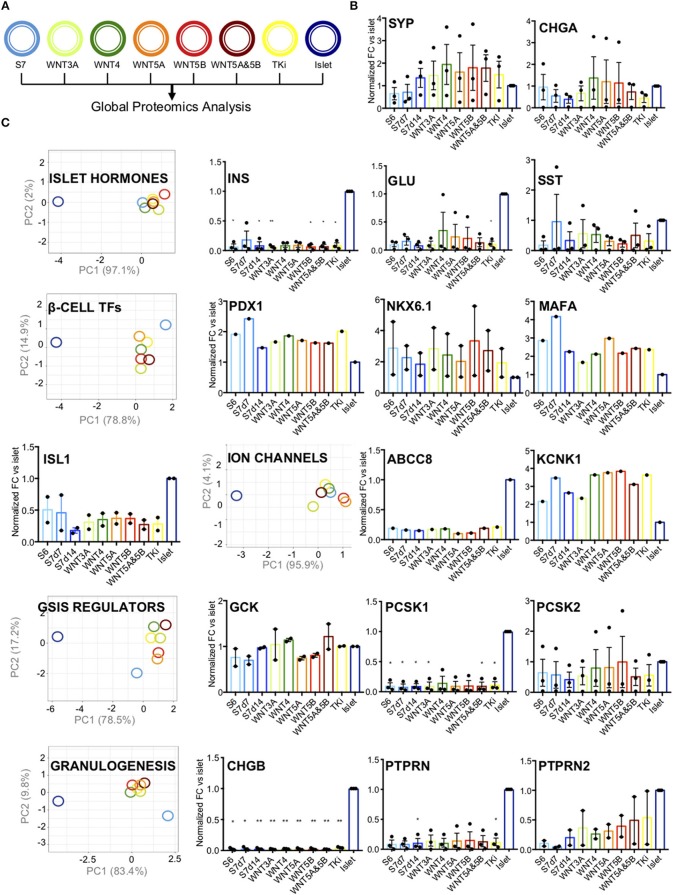
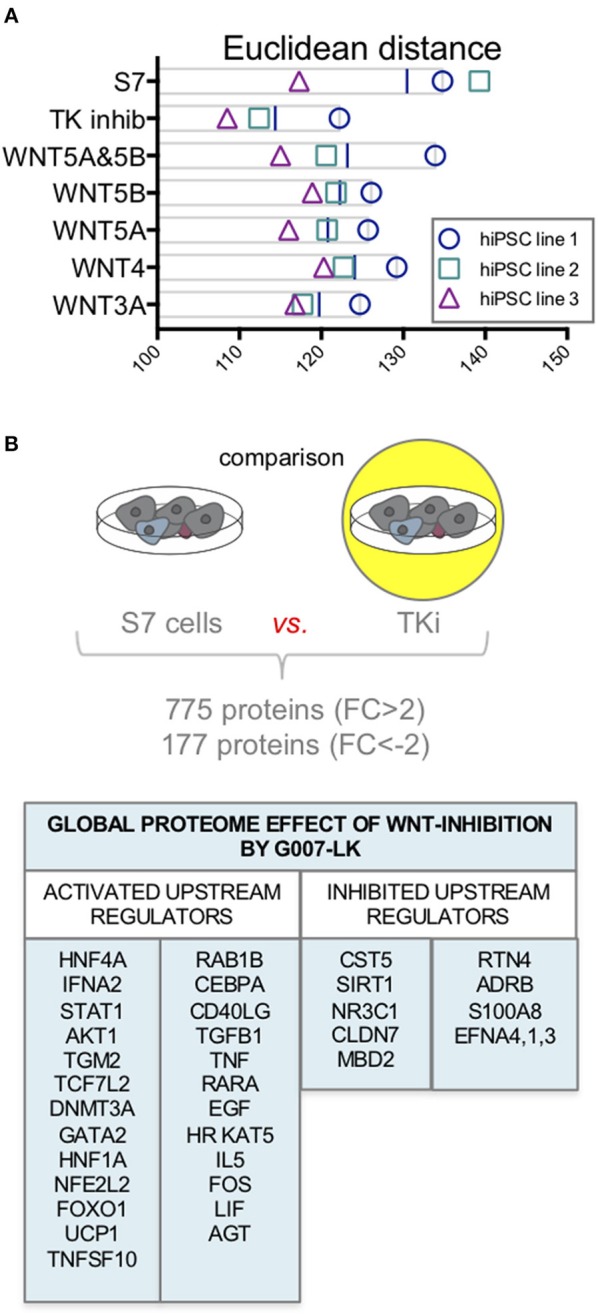
Current published protocols for targeted differentiation of human stem cells toward pancreatic β-cells fail to deliver sufficiently mature cells with functional properties comparable to human islet β-cells. We aimed to assess whether Wnt-modulation could promote the final protocol stages of β-cell maturation, building our hypothesis on our previous findings of Wnt activation in immature hiPSC-derived stage 7 (S7) cells compared to adult human islets and with recent data reporting a link between Wnt/PCP and in vitro β-cell maturation. In this study, we stimulated canonical and non-canonical Wnt signaling in hiPSC-derived S7 cells using syntetic proteins including WNT3A, WNT4, WNT5A and WNT5B, and we inhibited endogenous Wnt signaling with the Tankyrase inhibitor G007-LK (TKi). Whereas neither canonical nor non-canonical Wnt stimulation alone was able to mature hiPSC-derived S7 cells, WNT-inhibition with TKi increased the fraction of monohormonal cells and global proteomics of TKi-treated S7 cells showed a proteomic signature more similar to adult human islets, suggesting that inhibition of endogenous Wnt contributes toward final β-cell maturation.
Keywords: TMT11-plex; Wnt signaling pathway; adult human islets; human induced pluripotent stem cell; in vitro maturation; proteomics; tankyrase inhibition; β-like cells.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials