

Reprogrammed marrow adipocytes contribute to myeloma-induced bone disease
- PMID: 31142679
- PMCID: PMC6999853
- DOI: 10.1126/scitranslmed.aau9087
Reprogrammed marrow adipocytes contribute to myeloma-induced bone disease
Abstract
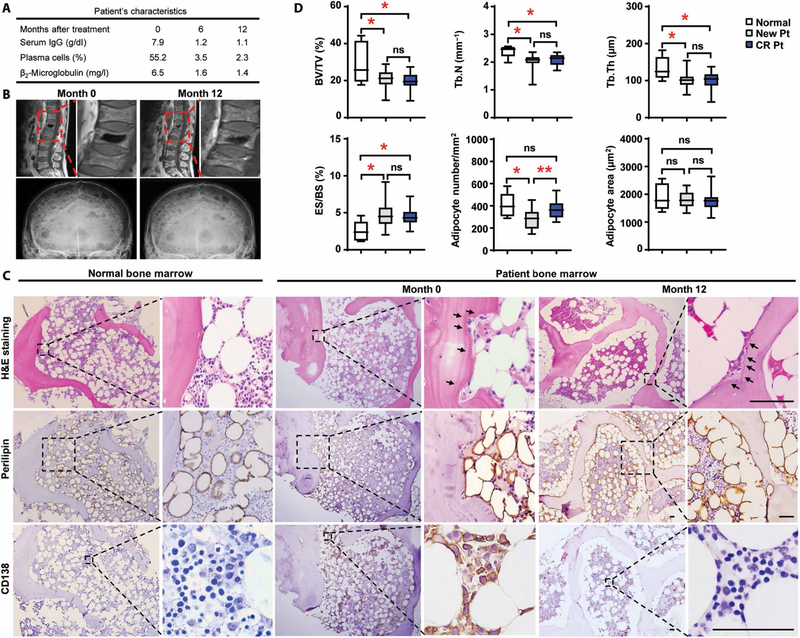
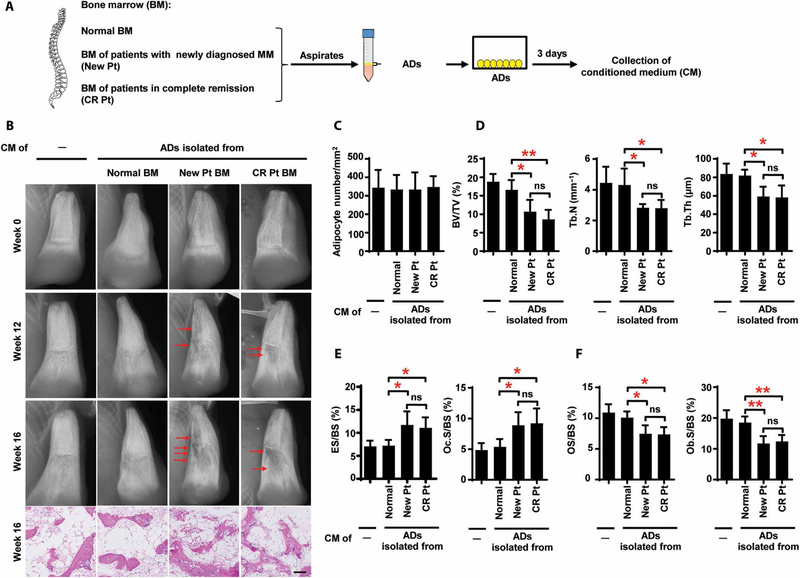
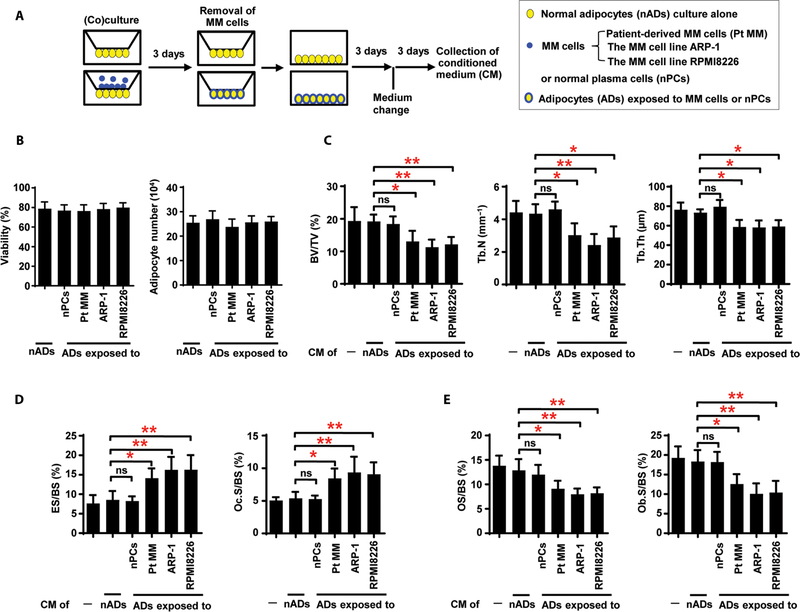
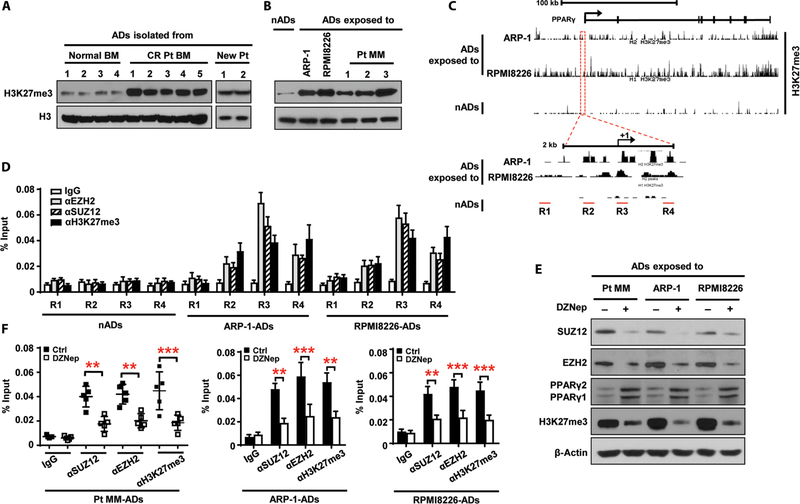
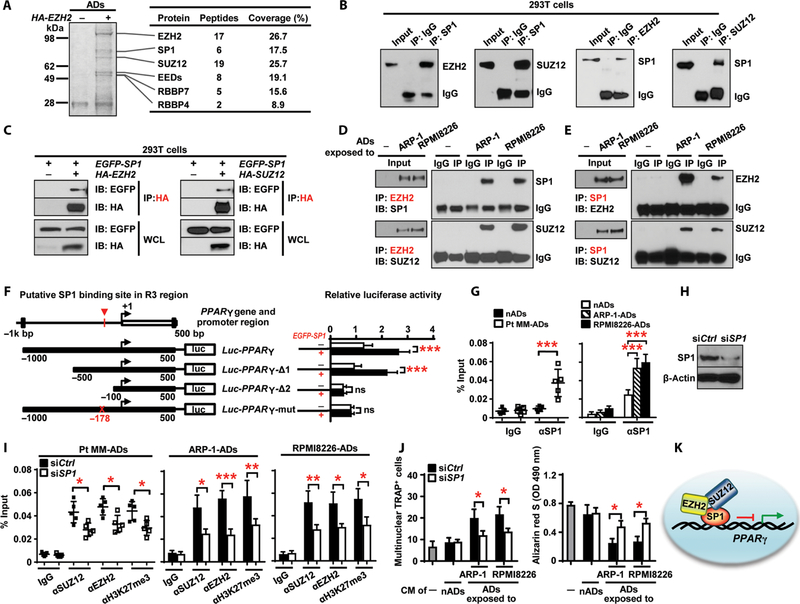
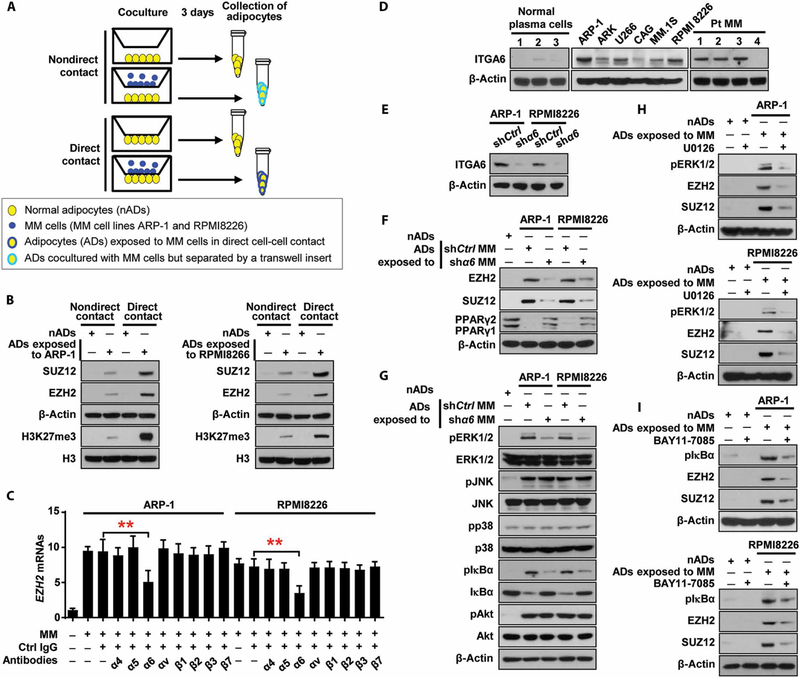
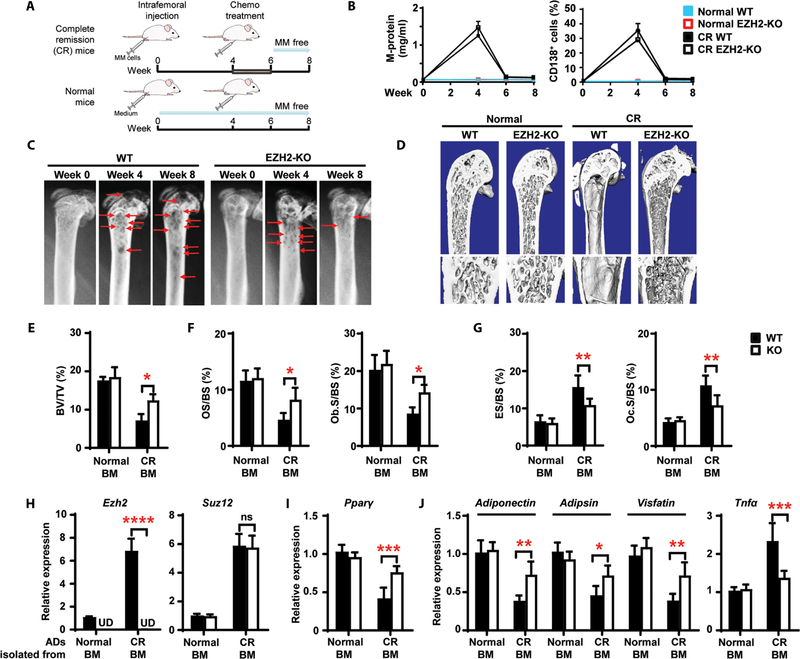
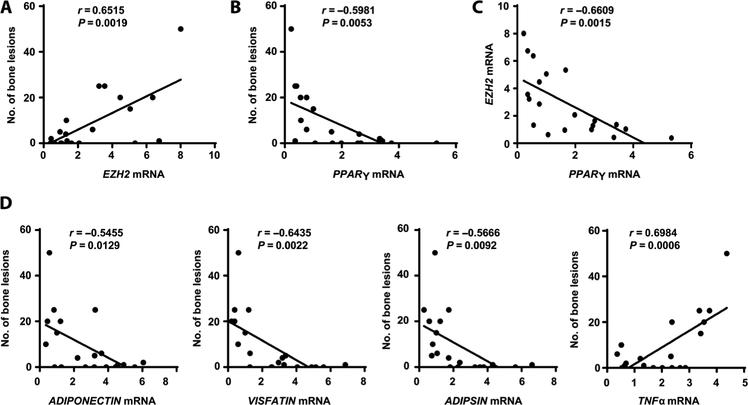
Osteolytic lesions in multiple myeloma are caused by osteoclast-mediated bone resorption and reduced bone formation. A unique feature of myeloma is a failure of bone healing after successful treatment. We observed adipocytes on trabecular bone near the resorbed area in successfully treated patients. Normal marrow adipocytes, when cocultured with myeloma cells, were reprogrammed and produced adipokines that activate osteoclastogenesis and suppress osteoblastogenesis. These adipocytes have reduced expression of peroxisome proliferator-activated receptor γ (PPARγ) mediated by recruitment of polycomb repressive complex 2 (PRC2), which modifies PPARγ promoter methylation at trimethyl lysine-27 histone H3. We confirmed the importance of methylation in the PPARγ promoter by demonstrating that adipocyte-specific knockout of EZH2, a member of the PRC2, prevents adipocyte reprogramming and reverses bone changes in a mouse model. We validated the strong correlation between the frequency of bone lesions and the expression of EZH2 in marrow adipocytes from patients in remission. These results define a role for adipocytes in genesis of myeloma-associated bone disease and that reversal of adipocyte reprogramming has therapeutic implications.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
New role for adipocytes in tumour-associated bone disease.Nat Rev Endocrinol. 2019 Aug;15(8):439. doi: 10.1038/s41574-019-0231-5. Nat Rev Endocrinol. 2019. PMID: 31227813 No abstract available.
References
-
- Palumbo A, Anderson K, Multiple myeloma. N. Engl. J. Med 364, 1046–1060 (2011). - PubMed
-
- Bruzzaniti A, Baron R, Molecular regulation of osteoclast activity. Rev. Endocr. Metab. Disord 7, 123–139 (2006). - PubMed
-
- Tian E, Zhan F, Walker R, Rasmussen E, Ma Y, Barlogie B, Shaughnessy JD Jr., The role of the Wnt-signaling antagonist DKK1 in the development of osteolytic lesions in multiple myeloma. N. Engl. J. Med 349, 2483–2494 (2003). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
