

Pushing the envelope: LPS modifications and their consequences
- PMID: 31142822
- PMCID: PMC6913091
- DOI: 10.1038/s41579-019-0201-x
Pushing the envelope: LPS modifications and their consequences
Abstract
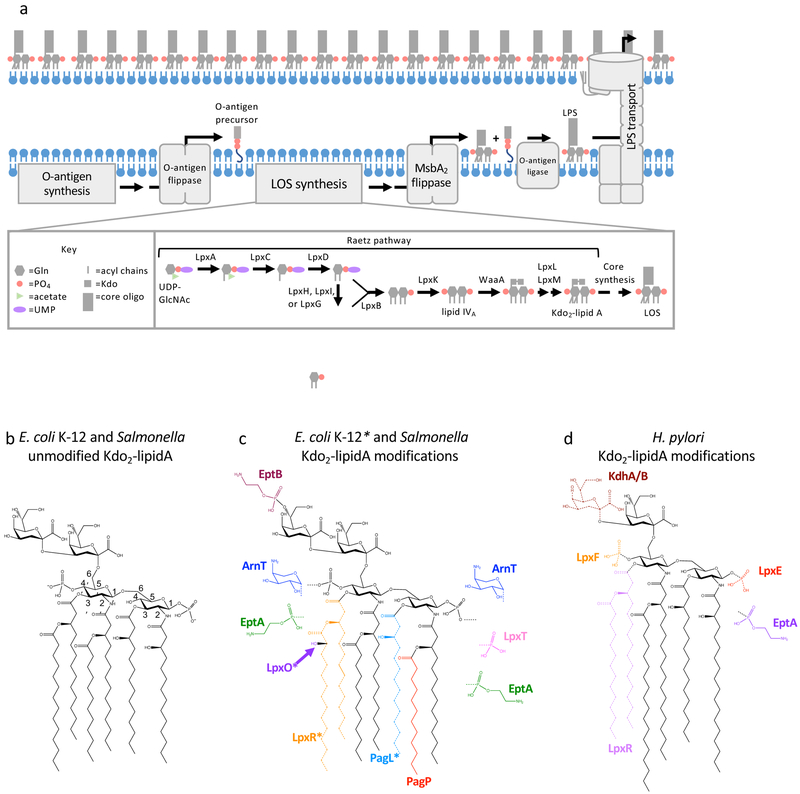
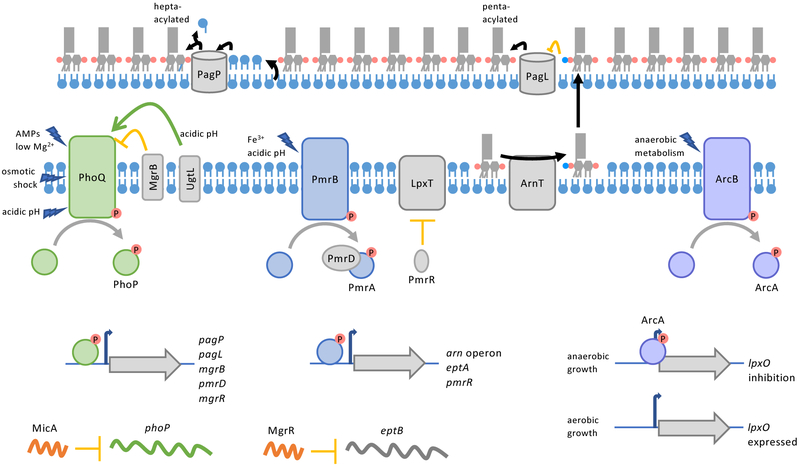
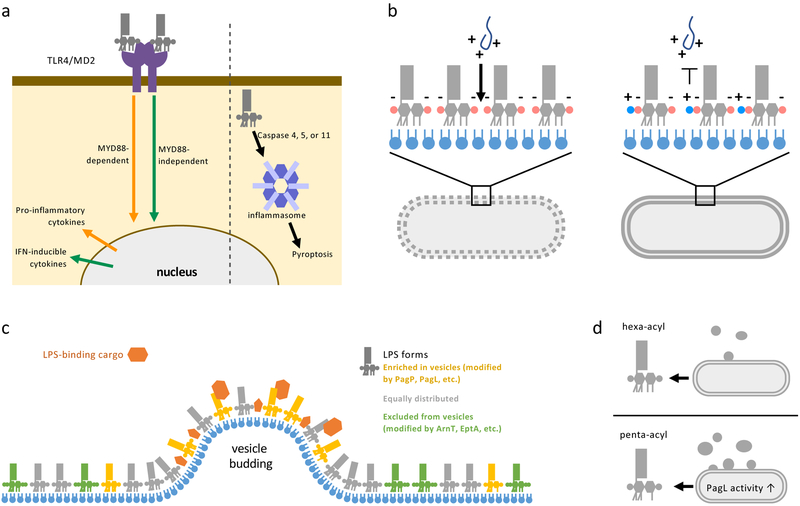
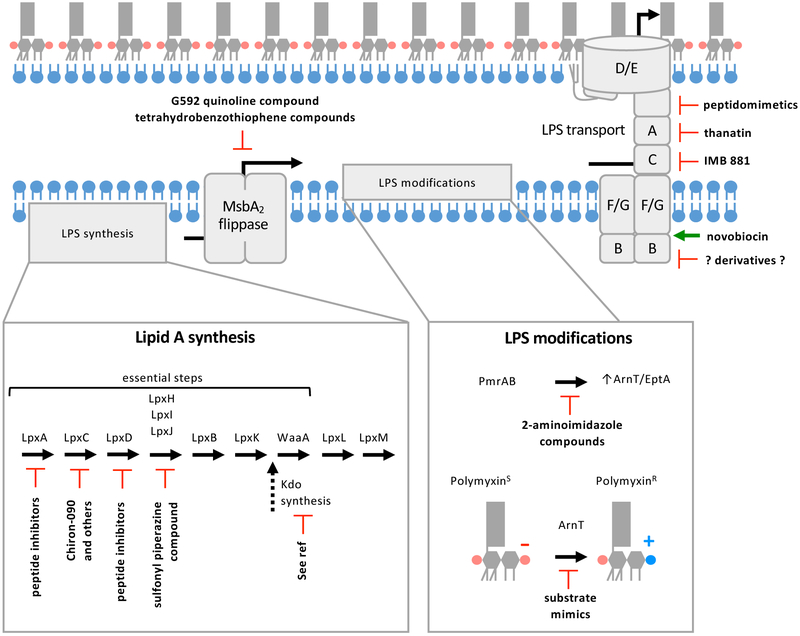
The defining feature of the Gram-negative cell envelope is the presence of two cellular membranes, with the specialized glycolipid lipopolysaccharide (LPS) exclusively found on the surface of the outer membrane. The surface layer of LPS contributes to the stringent permeability properties of the outer membrane, which is particularly resistant to permeation of many toxic compounds, including antibiotics. As a common surface antigen, LPS is recognized by host immune cells, which mount defences to clear pathogenic bacteria. To alter properties of the outer membrane or evade the host immune response, Gram-negative bacteria chemically modify LPS in a wide variety of ways. Here, we review key features and physiological consequences of LPS biogenesis and modifications.
Figures
References
-
- Muhlradt PF & Golecki JR Asymmetrical distribution and artifactual reorientation of lipopolysaccharide in the outer membrane bilayer of Salmonella typhimurium. European journal of biochemistry 51, 343–352 (1975). - PubMed
-
- Muhlradt PF, Menzel J, Golecki JR & Speth V Outer membrane of salmonella. Sites of export of newly synthesised lipopolysaccharide on the bacterial surface. European journal of biochemistry 35, 471–481 (1973). - PubMed
-
- Osborn MJ, Gander JE & Parisi E Mechanism of assembly of the outer membrane of Salmonella typhimurium. Site of synthesis of lipopolysaccharide. The Journal of biological chemistry 247, 3973–3986 (1972). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
