

Mouse Models of Heterologous Flavivirus Immunity: A Role for Cross-Reactive T Cells
- PMID: 31143185
- PMCID: PMC6520664
- DOI: 10.3389/fimmu.2019.01045
Mouse Models of Heterologous Flavivirus Immunity: A Role for Cross-Reactive T Cells
Abstract
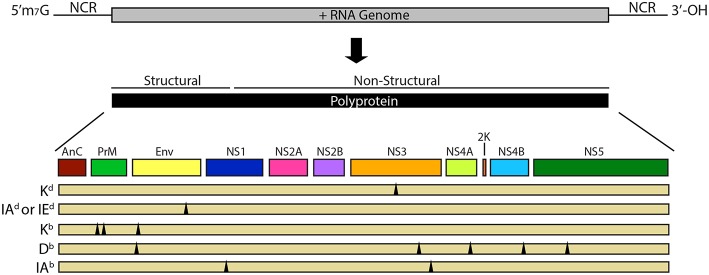
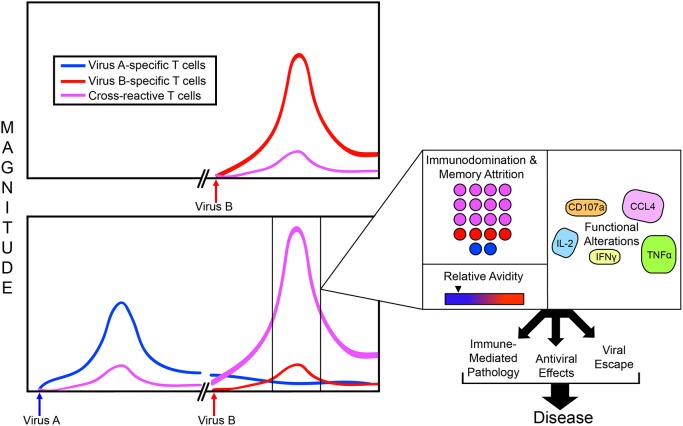
Most of the world is at risk of being infected with a flavivirus such as dengue virus, West Nile virus, yellow fever virus, Japanese encephalitis virus, tick-borne encephalitis virus, and Zika virus, significantly impacting millions of lives. Importantly, many of these genetically similar viruses co-circulate within the same geographic regions, making it likely for individuals living in areas of high flavivirus endemicity to be infected with multiple flaviviruses during their lifetime. Following a flavivirus infection, a robust virus-specific T cell response is generated and the memory recall of this response has been demonstrated to provide long-lasting immunity, protecting against reinfection with the same pathogen. However, multiple studies have shown that this flavivirus specific T cell response can be cross-reactive and active during heterologous flavivirus infection, leading to the question: How does immunity to one flavivirus shape immunity to the next, and how does this impact disease? It has been proposed that in some cases unfavorable disease outcomes may be caused by lower avidity cross-reactive memory T cells generated during a primary flavivirus infection that preferentially expand during a secondary heterologous infection and function sub optimally against the new pathogen. While in other cases, these cross-reactive cells still have the potential to facilitate cross-protection. In this review, we focus on cross-reactive T cell responses to flaviviruses and the concepts and consequences of T cell cross-reactivity, with particular emphasis linking data generated using murine models to our new understanding of disease outcomes following heterologous flavivirus infection.
Keywords: T cell cross-reactivity; Zika; dengue; flavivirus; heterologous immunity; original antigenic sin.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources