

PETISCO is a novel protein complex required for 21U RNA biogenesis and embryonic viability
- PMID: 31147388
- PMCID: PMC6601512
- DOI: 10.1101/gad.322446.118
PETISCO is a novel protein complex required for 21U RNA biogenesis and embryonic viability
Abstract
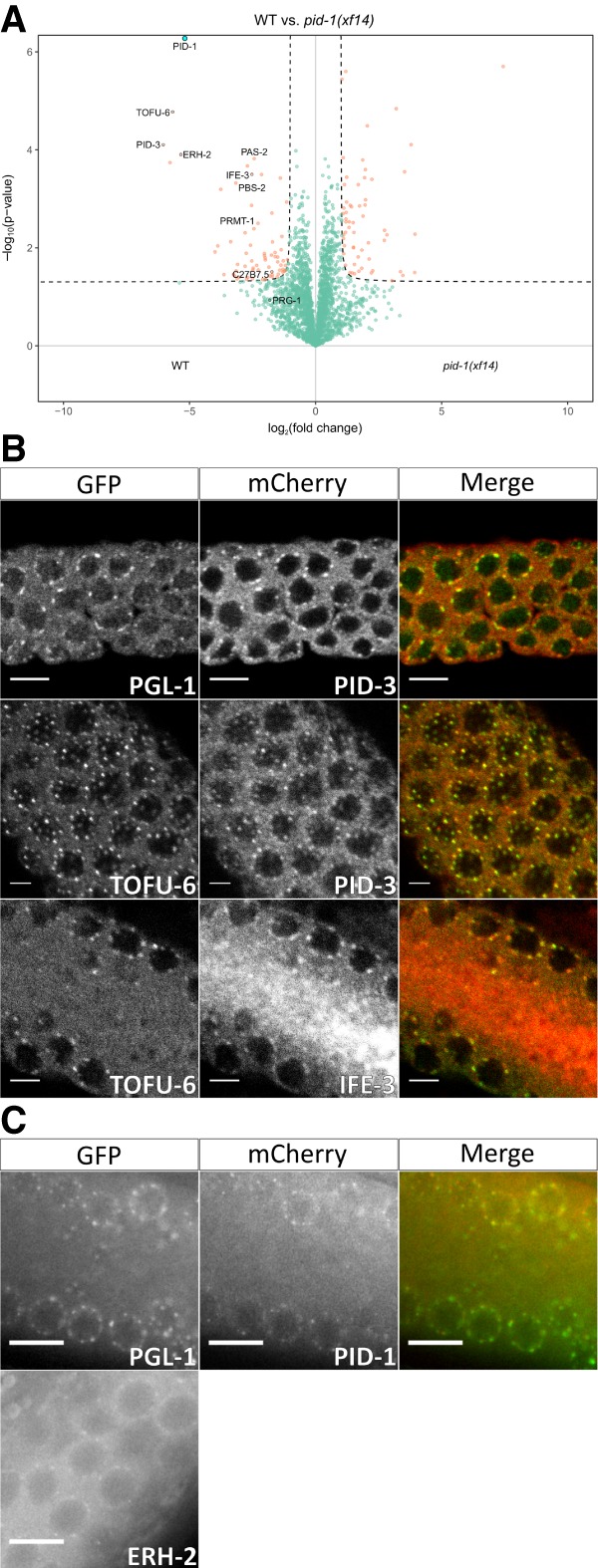
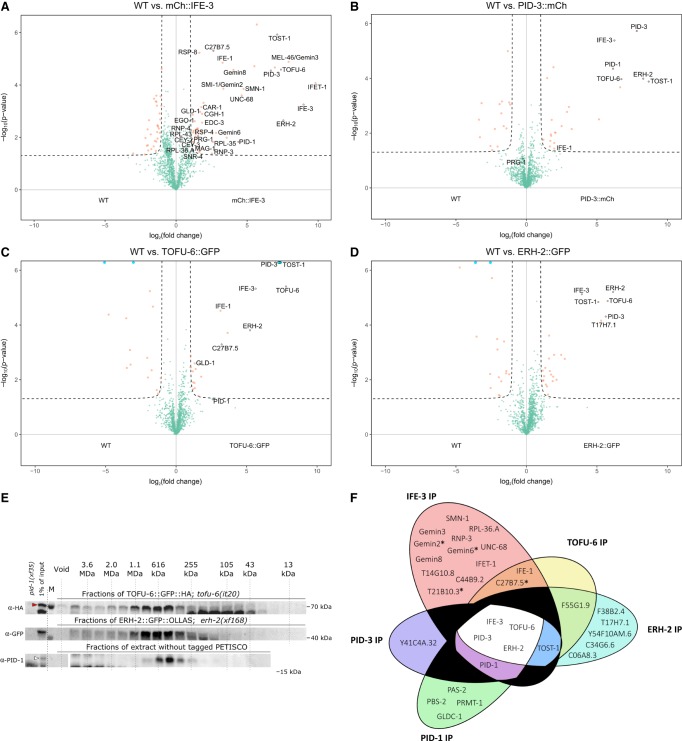
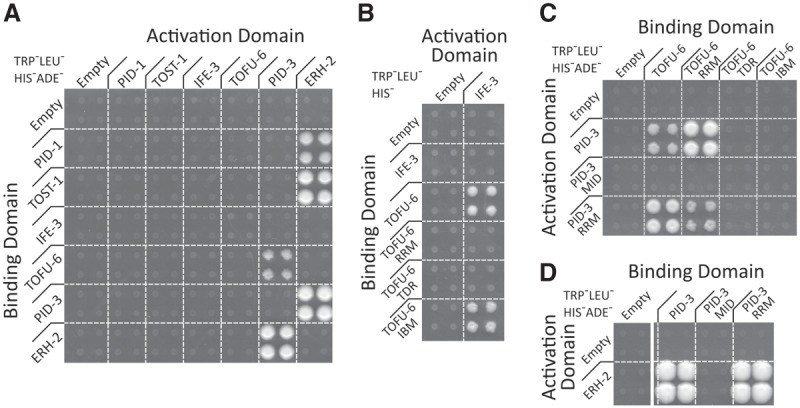
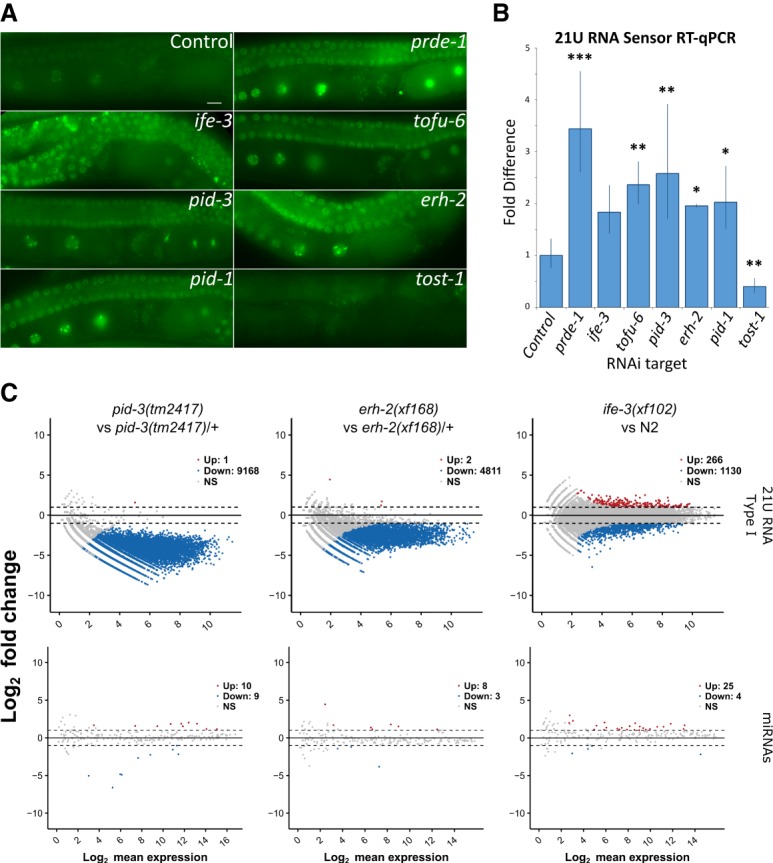
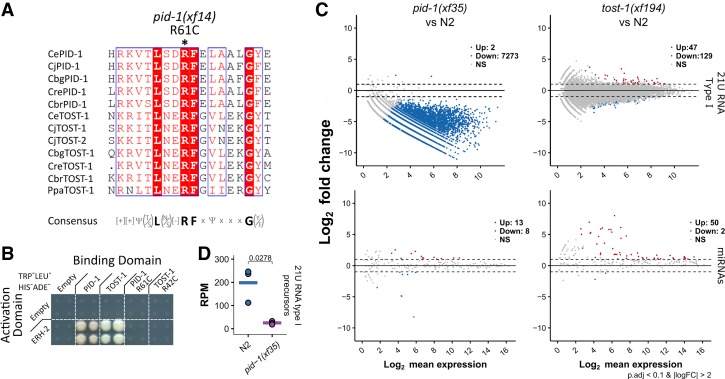
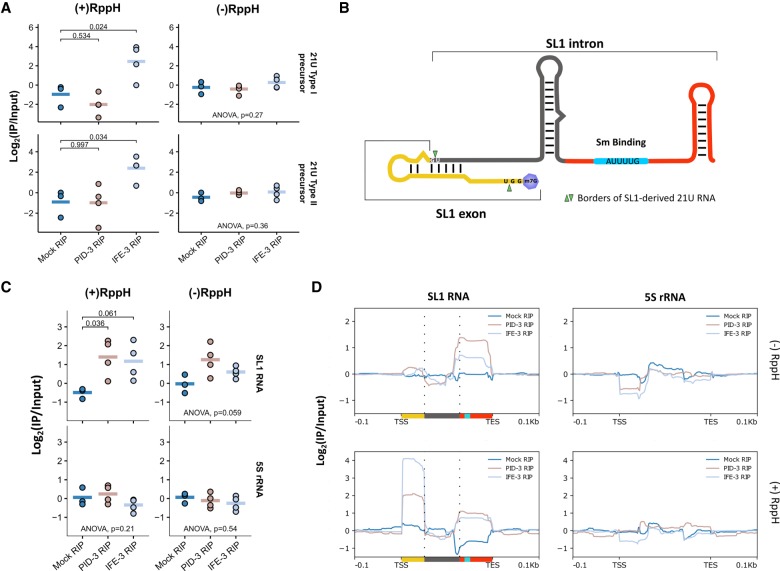
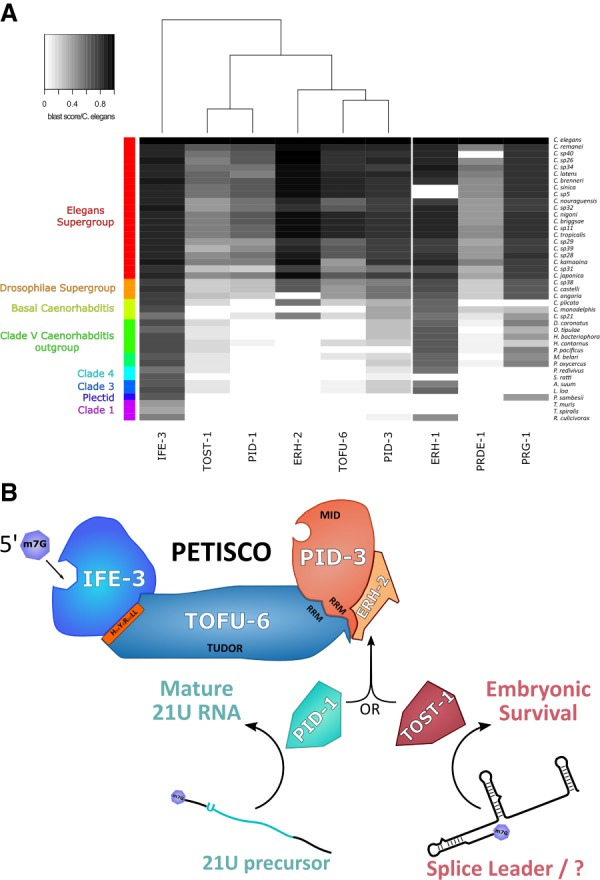
Piwi proteins are important for germ cell development in most animals. These proteins are guided to specific targets by small guide RNAs, referred to as piRNAs or 21U RNAs in Caenorhabditis elegans In this organism, even though genetic screens have uncovered 21U RNA biogenesis factors, little is known about how these factors interact or what they do. Based on the previously identified 21U biogenesis factor PID-1 (piRNA-induced silencing-defective 1), we here define a novel protein complex, PETISCO (PID-3, ERH-2, TOFU-6, and IFE-3 small RNA complex), that is required for 21U RNA biogenesis. PETISCO contains both potential 5' cap and 5' phosphate RNA-binding domains and interacts with capped 21U precursor RNA. We resolved the architecture of PETISCO and revealed a second function for PETISCO in embryonic development. This essential function of PETISCO is mediated not by PID-1 but by the novel protein TOST-1 (twenty-one U pathway antagonist). In contrast, TOST-1 is not essential for 21U RNA biogenesis. Both PID-1 and TOST-1 interact directly with ERH-2 using a conserved sequence motif. Finally, our data suggest a role for TOST-1:PETISCO in SL1 homeostasis in the early embryo. Our work describes a key complex for 21U RNA processing in C. elegans and strengthens the view that 21U RNA biogenesis is built on an snRNA-related pathway.
Keywords: 21U RNA; C. elegans; IFE-3; MID domain; PID-1; PID-3; PRG-1; TOFU-6; enhancer of rudimentary; piRNA.
© 2019 Cordeiro Rodrigues et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous