

Structure of the super-elongation complex subunit AFF4 C-terminal homology domain reveals requirements for AFF homo- and heterodimerization
- PMID: 31147444
- PMCID: PMC6615702
- DOI: 10.1074/jbc.RA119.008577
Structure of the super-elongation complex subunit AFF4 C-terminal homology domain reveals requirements for AFF homo- and heterodimerization
Abstract
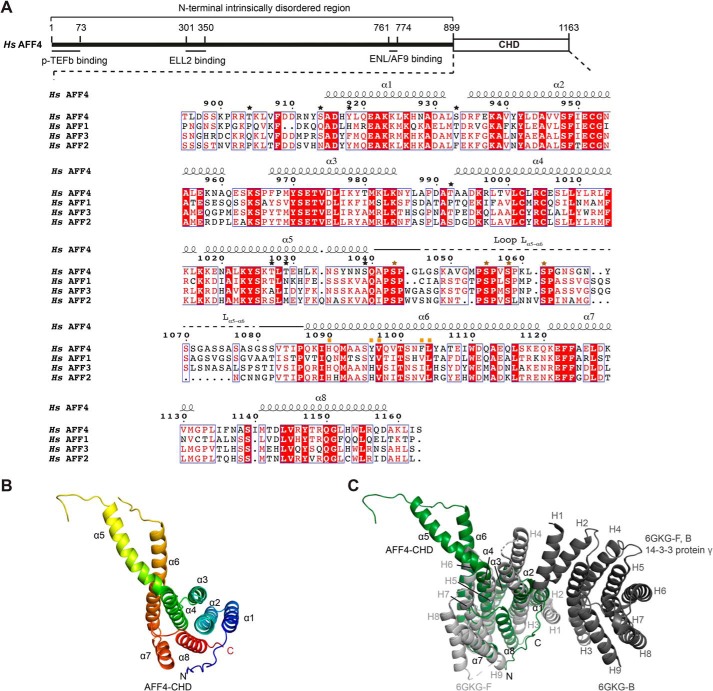
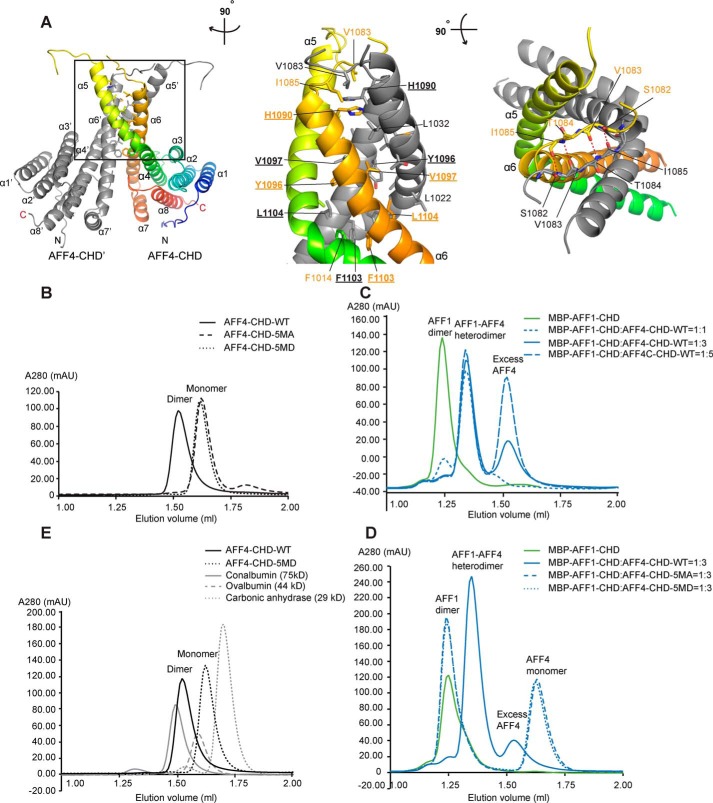
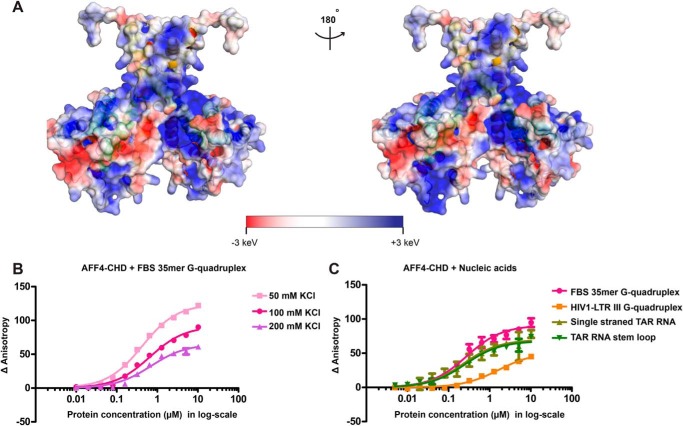
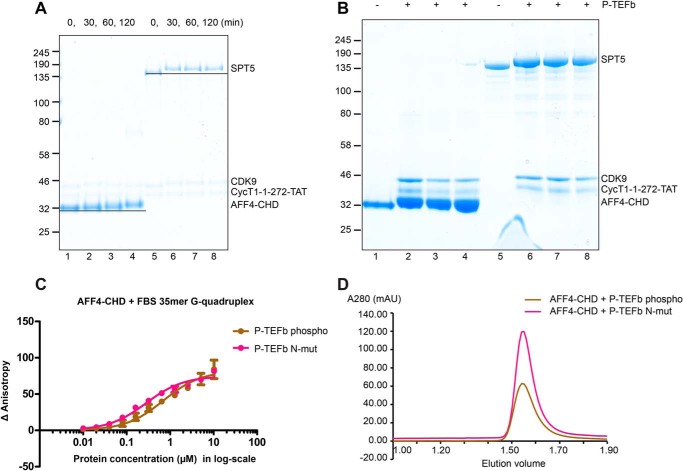
AF4/FMR2 family member 4 (AFF4) is the scaffold protein of the multisubunit super-elongation complex, which plays key roles in the release of RNA polymerase II from promoter-proximal pausing and in the transactivation of HIV-1 transcription. AFF4 consists of an intrinsically disordered N-terminal region that interacts with other super-elongation complex subunits and a C-terminal homology domain (CHD) that is conserved among AF4/FMR2 family proteins, including AFF1, AFF2, AFF3, and AFF4. Here, we solved the X-ray crystal structure of the CHD in human AFF4 (AFF4-CHD) to 2.2 Å resolution and characterized its biochemical properties. The structure disclosed that AFF4-CHD folds into a novel domain that consists of eight helices and is distantly related to tetratrico peptide repeat motifs. Our analyses further revealed that AFF4-CHD mediates the formation of an AFF4 homodimer or an AFF1-AFF4 heterodimer. Results from fluorescence anisotropy experiments suggested that AFF4-CHD interacts with both RNA and DNA in vitro Furthermore, we identified a surface loop region in AFF4-CHD as a substrate for the P-TEFb kinase cyclin-dependent kinase 9, which triggers release of polymerase II from promoter-proximal pausing sites. In conclusion, the AFF-CHD structure and biochemical analyses reported here reveal the molecular basis for the homo- and heterodimerization of AFF proteins and implicate the AFF4-CHD in nucleic acid interactions. The high conservation of the CHD among several other proteins suggests that our results are also relevant for understanding other CHD-containing proteins and their dimerization behavior.
Keywords: AF4/FMR2 family member 4 (AFF4); C-terminal homology domain (CHD); RNA polymerase II; X-ray crystallography; dimerization; gene regulation; phosphorylation; super elongation complex; transcription elongation factor.
© 2019 Chen and Cramer.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
