

Building bridges to move recombination complexes
- PMID: 31147459
- PMCID: PMC6589682
- DOI: 10.1073/pnas.1901237116
Building bridges to move recombination complexes
Abstract
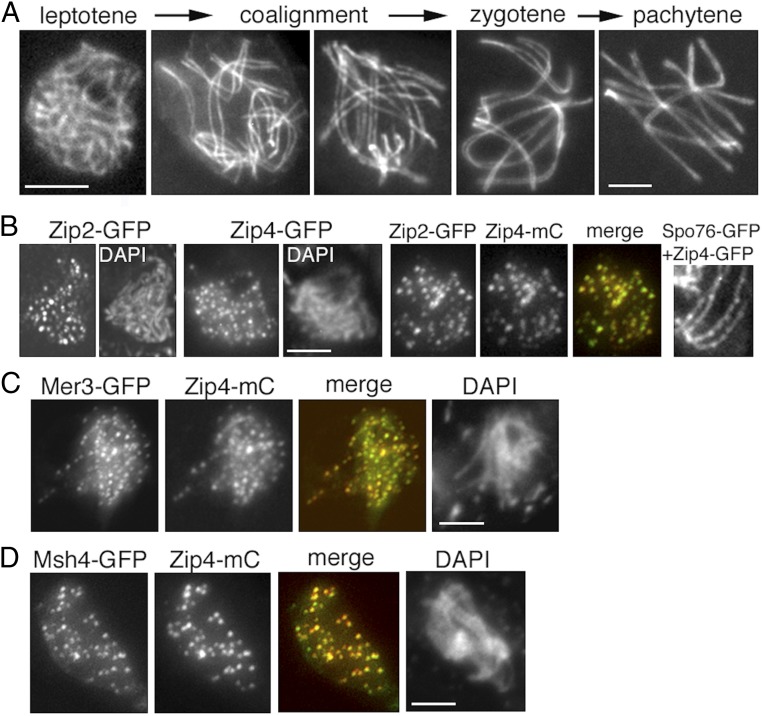
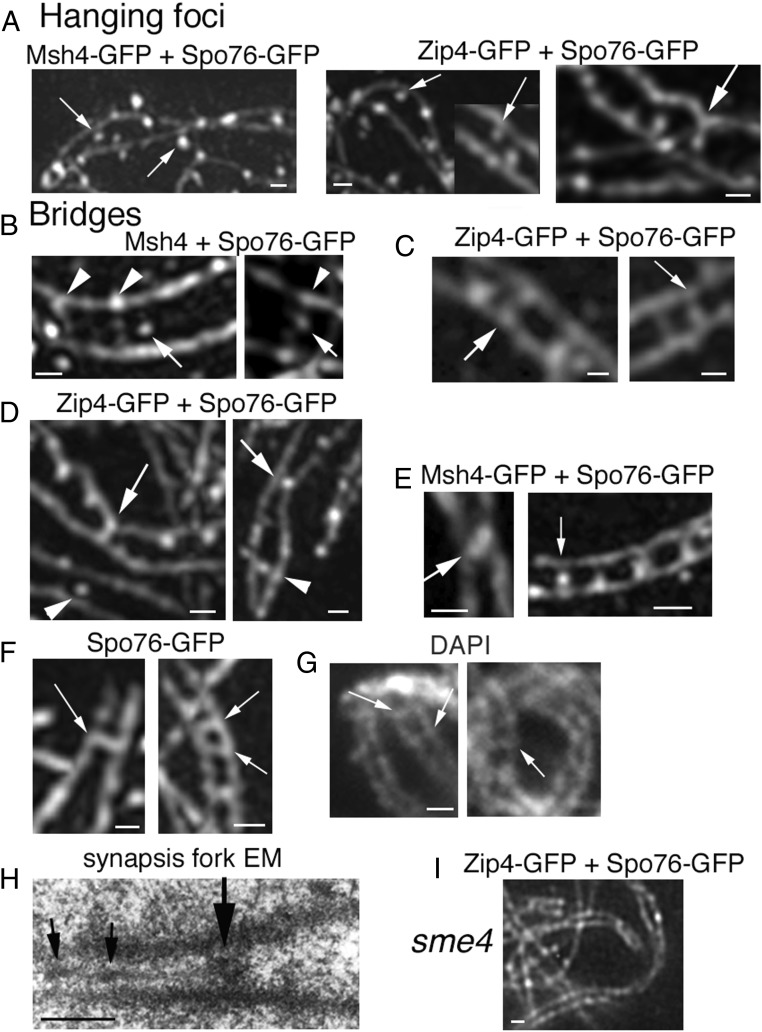
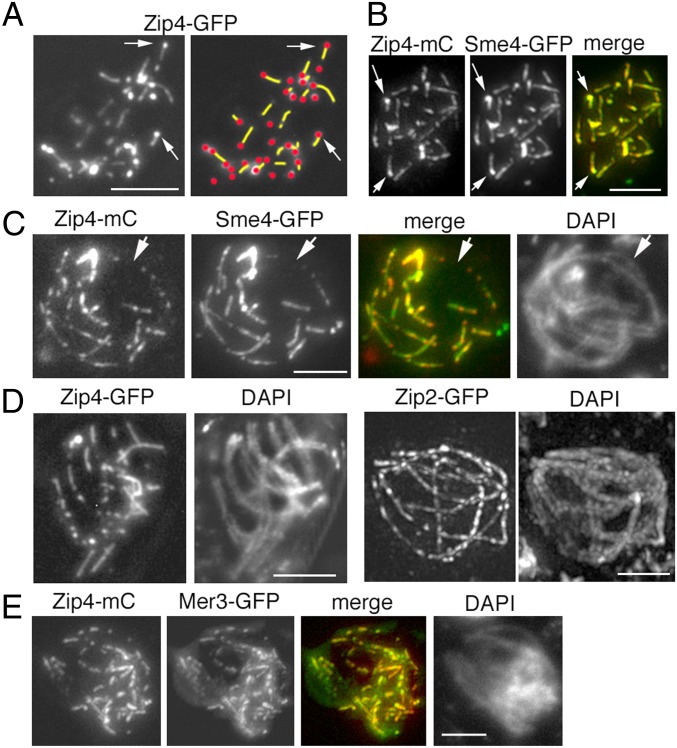
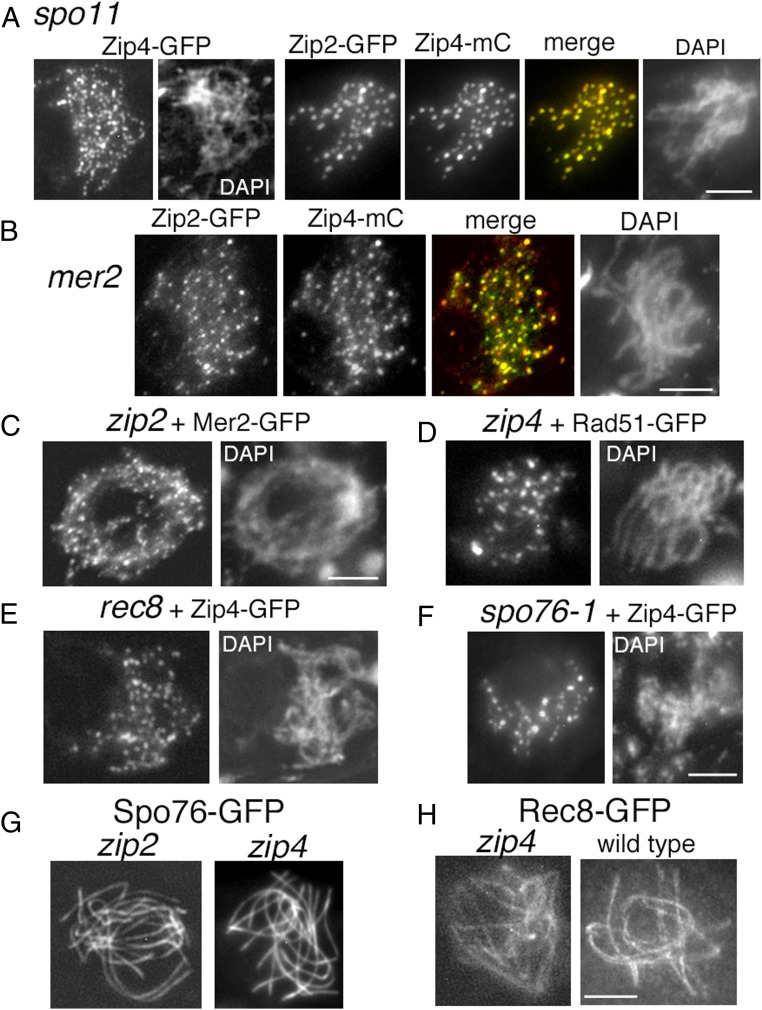
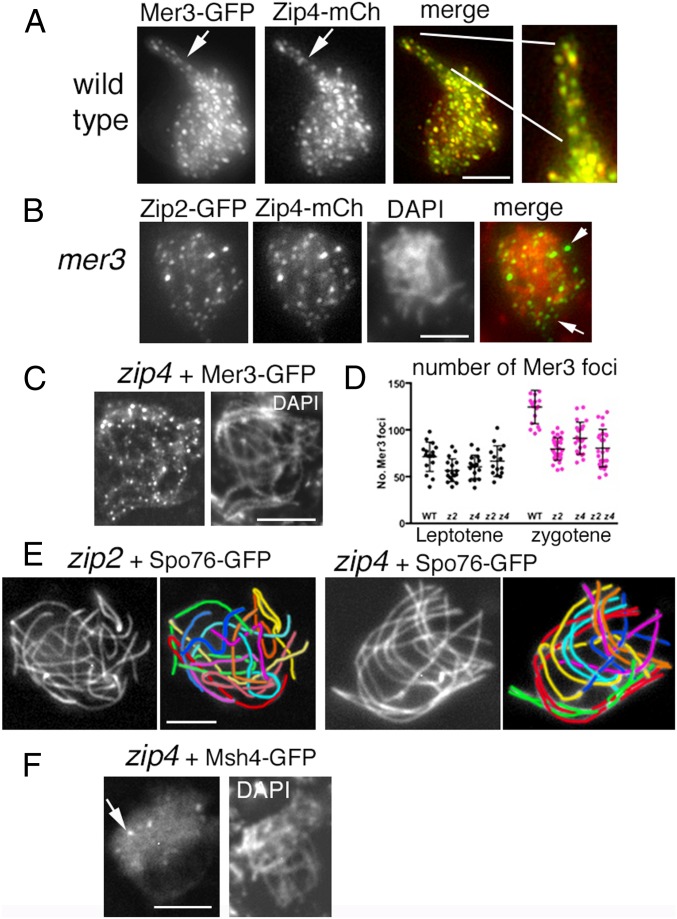
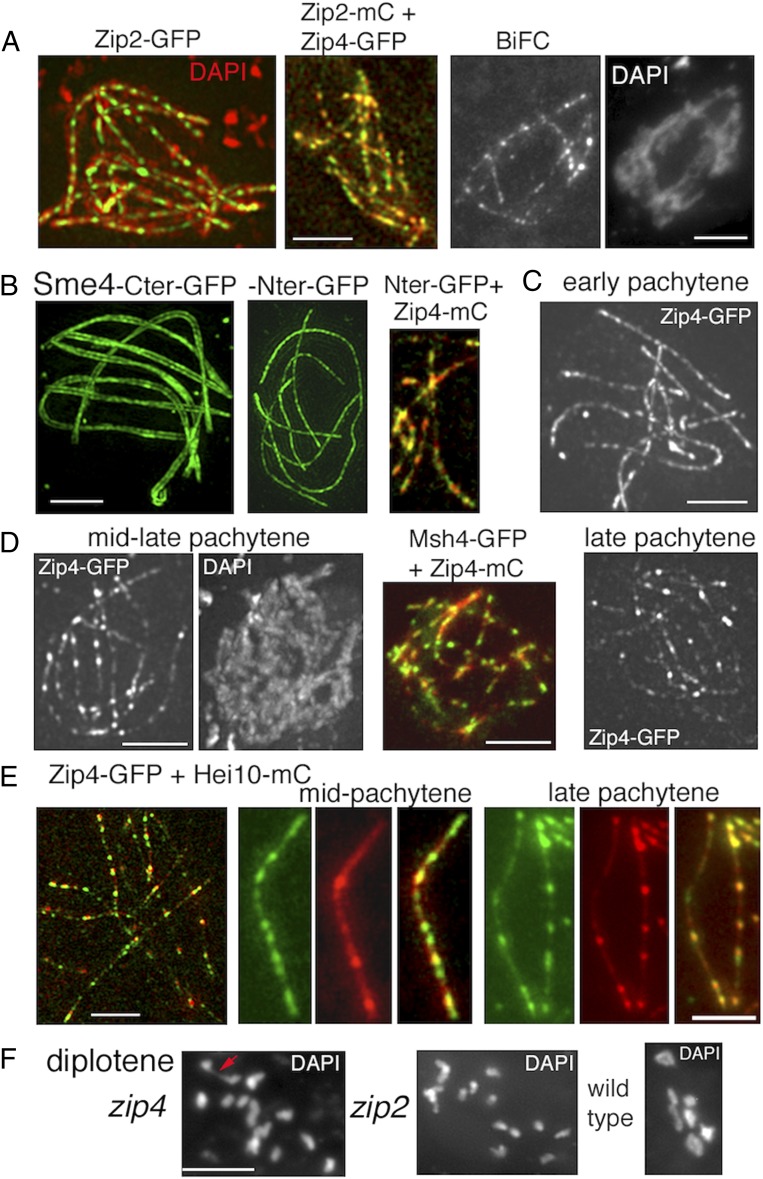
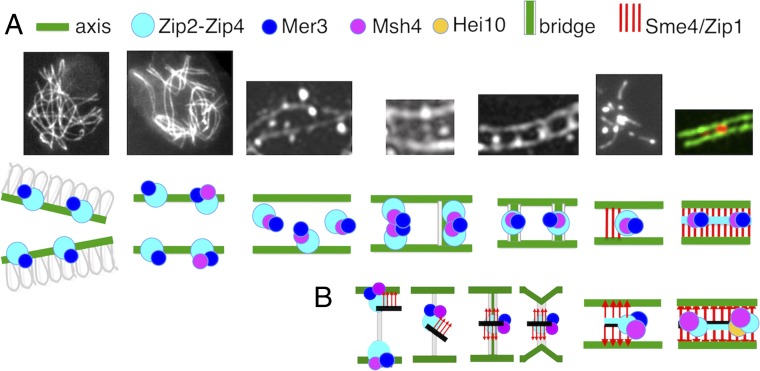
A central feature of meiosis is pairing of homologous chromosomes, which occurs in two stages: coalignment of axes followed by installation of the synaptonemal complex (SC). Concomitantly, recombination complexes reposition from on-axis association to the SC central region. We show here that, in the fungus Sordaria macrospora, this critical transition is mediated by robust interaxis bridges that contain an axis component (Spo76/Pds5), DNA, plus colocalizing Mer3/Msh4 recombination proteins and the Zip2-Zip4 mediator complex. Mer3-Msh4-Zip2-Zip4 colocalizing foci are first released from their tight axis association, dependent on the SC transverse-filament protein Sme4/Zip1, before moving to bridges and thus to a between-axis position. Ensuing shortening of bridges and accompanying juxtaposition of axes to 100 nm enables installation of SC central elements at sites of between-axis Mer3-Msh4-Zip2-Zip4 complexes. We show also that the Zip2-Zip4 complex has an intrinsic affinity for chromosome axes at early leptotene, where it localizes independently of recombination, but is dependent on Mer3. Then, later, Zip2-Zip4 has an intrinsic affinity for the SC central element, where it ultimately localizes to sites of crossover complexes at the end of pachytene. These and other findings suggest that the fundamental role of Zip2-Zip4 is to mediate the recombination/structure interface at all post-double-strand break stages. We propose that Zip2-Zip4 directly mediates a molecular handoff of Mer3-Msh4 complexes, from association with axis components to association with SC central components, at the bridge stage, and then directly mediates central region installation during SC nucleation.
Keywords: Zip2-Zip4; chromosome structure; interaxis bridges; meiotic recombination; synaptonemal complex.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
