

Curvature induction and membrane remodeling by FAM134B reticulon homology domain assist selective ER-phagy
- PMID: 31147549
- PMCID: PMC6542808
- DOI: 10.1038/s41467-019-10345-3
Curvature induction and membrane remodeling by FAM134B reticulon homology domain assist selective ER-phagy
Abstract
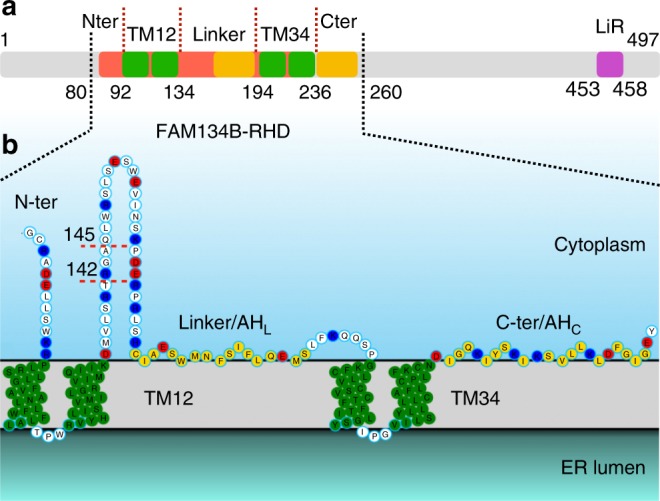
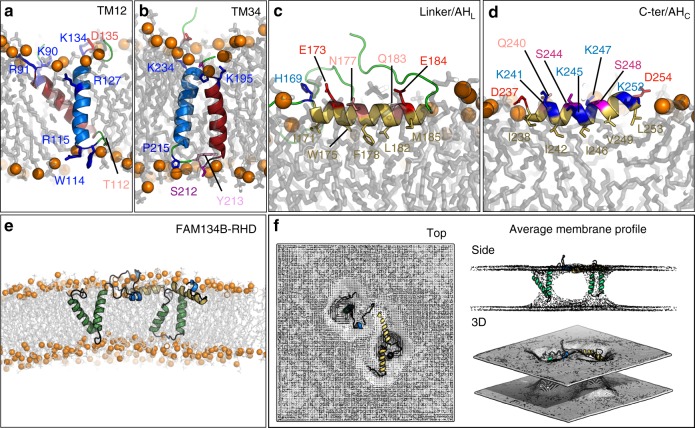
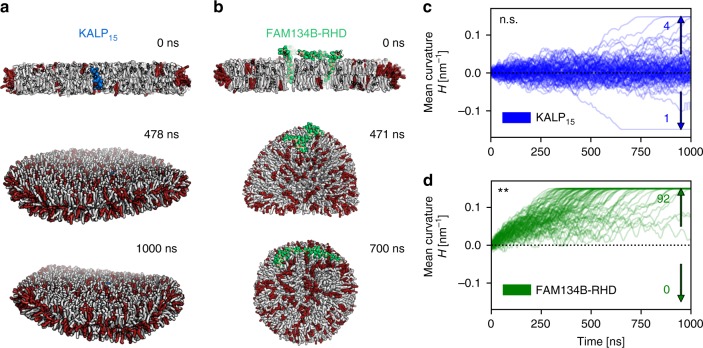
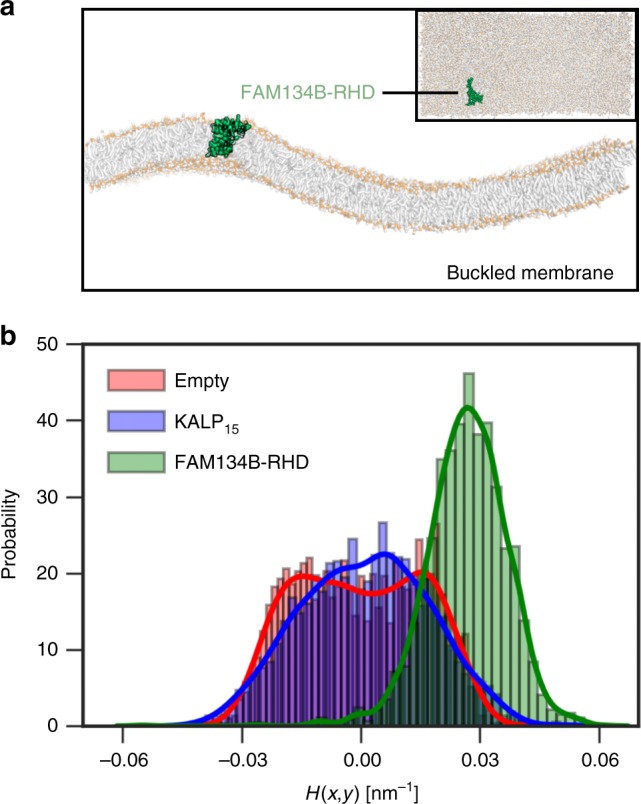
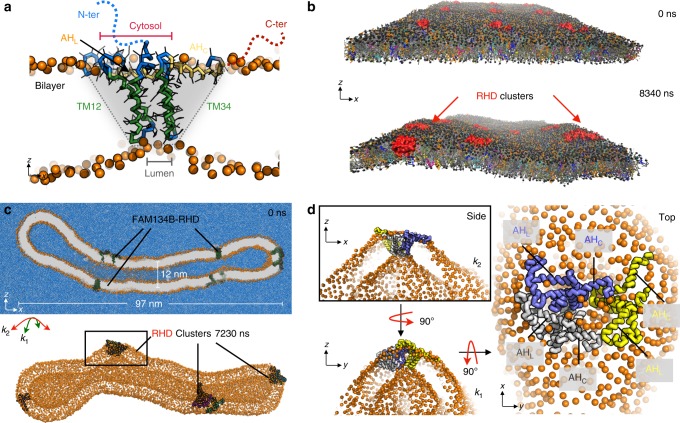
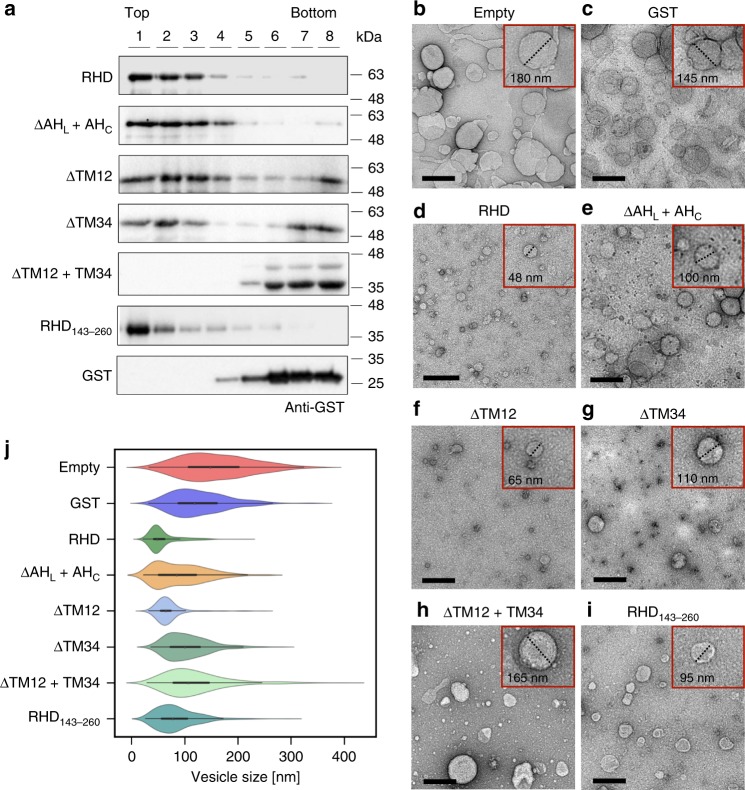
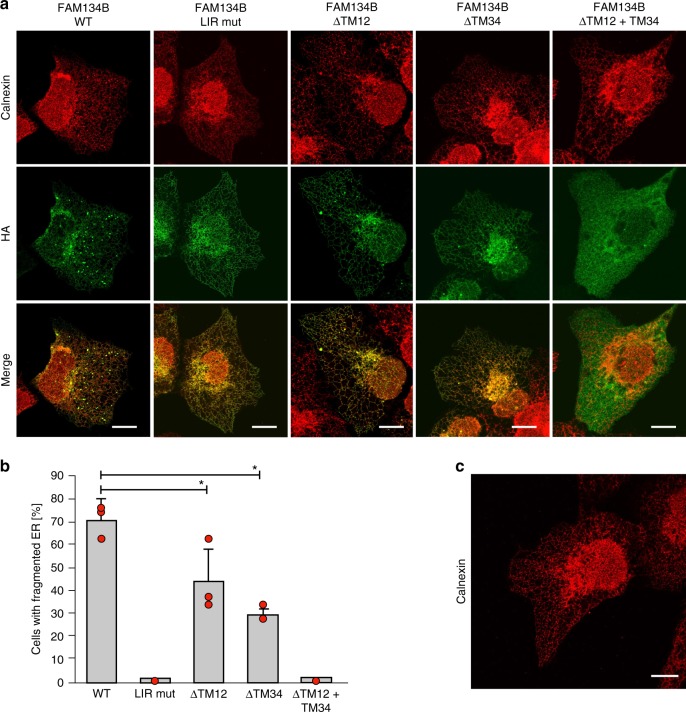
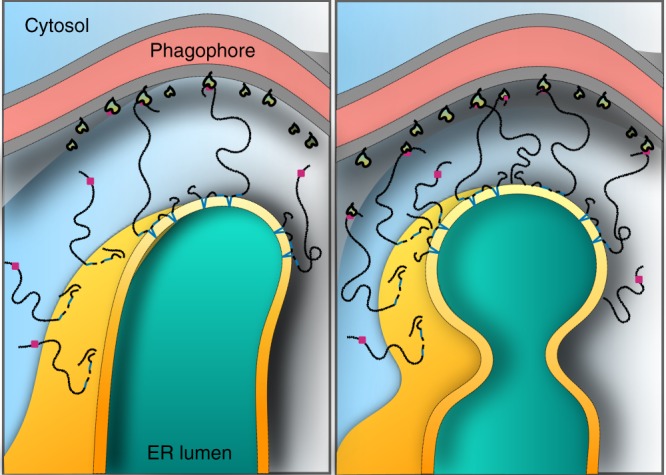
FAM134B/RETREG1 is a selective ER-phagy receptor that regulates the size and shape of the endoplasmic reticulum. The structure of its reticulon-homology domain (RHD), an element shared with other ER-shaping proteins, and the mechanism of membrane shaping remain poorly understood. Using molecular modeling and molecular dynamics (MD) simulations, we assemble a structural model for the RHD of FAM134B. Through MD simulations of FAM134B in flat and curved membranes, we relate the dynamic RHD structure with its two wedge-shaped transmembrane helical hairpins and two amphipathic helices to FAM134B functions in membrane-curvature induction and curvature-mediated protein sorting. FAM134B clustering, as expected to occur in autophagic puncta, amplifies the membrane-shaping effects. Electron microscopy of in vitro liposome remodeling experiments support the membrane remodeling functions of the different RHD structural elements. Disruption of the RHD structure affects selective autophagy flux and leads to disease states.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- Sofja Kovalevskaja Award/Alexander von Humboldt-Stiftung (Alexander von Humboldt Foundation)/International
- SFB 1177/Deutsche Forschungsgemeinschaft (German Research Foundation)/International
- 259130777/Deutsche Forschungsgemeinschaft (German Research Foundation)/International
- 742720 UbBAC,/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 European Research Council (H2020 Excellent Science - European Research Council)/International
LinkOut - more resources
Full Text Sources
