

The next generation of CRISPR-Cas technologies and applications
- PMID: 31147612
- PMCID: PMC7079207
- DOI: 10.1038/s41580-019-0131-5
The next generation of CRISPR-Cas technologies and applications
Abstract
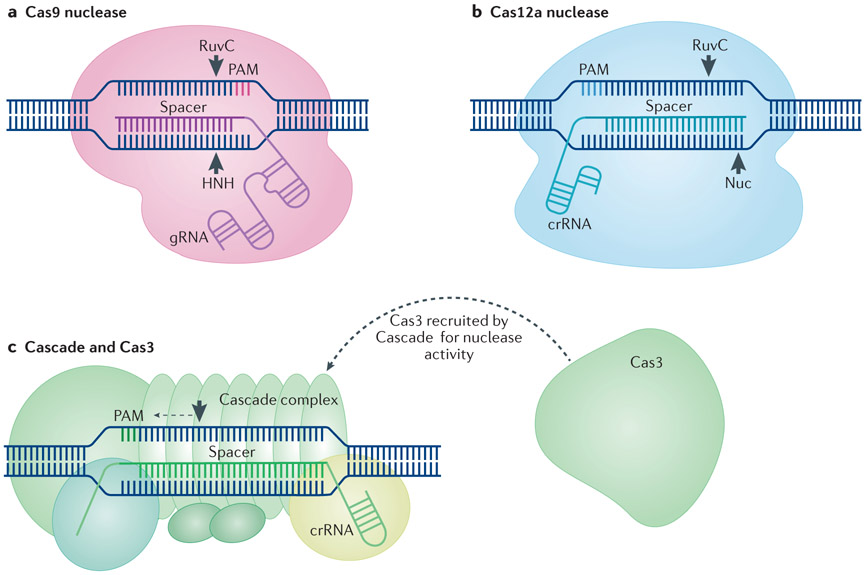
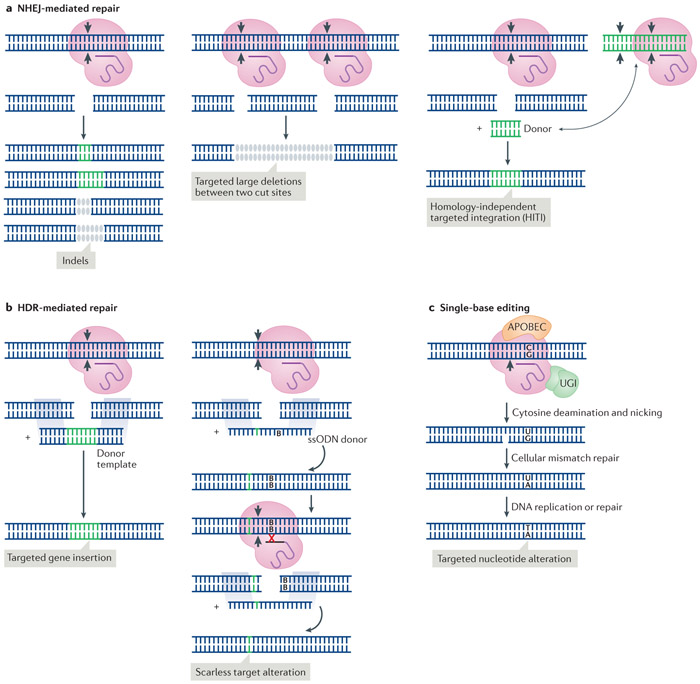
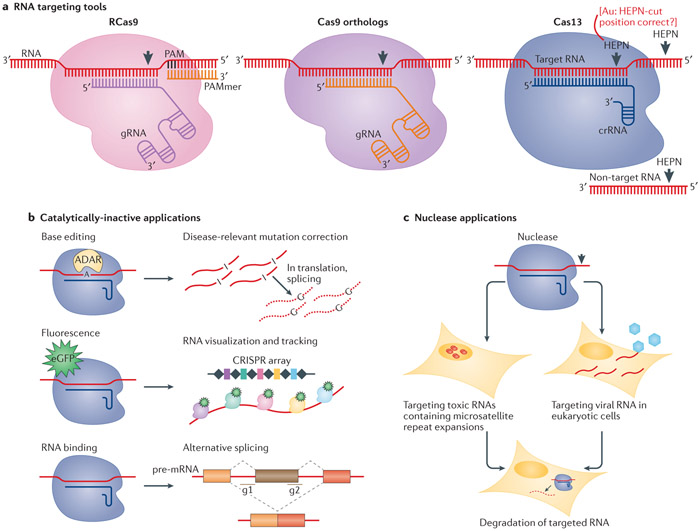
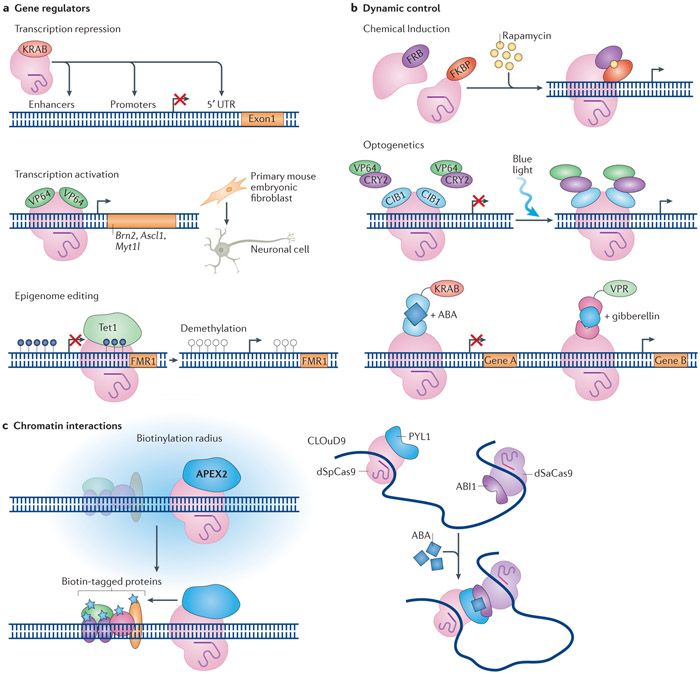
The prokaryote-derived CRISPR-Cas genome editing systems have transformed our ability to manipulate, detect, image and annotate specific DNA and RNA sequences in living cells of diverse species. The ease of use and robustness of this technology have revolutionized genome editing for research ranging from fundamental science to translational medicine. Initial successes have inspired efforts to discover new systems for targeting and manipulating nucleic acids, including those from Cas9, Cas12, Cascade and Cas13 orthologues. Genome editing by CRISPR-Cas can utilize non-homologous end joining and homology-directed repair for DNA repair, as well as single-base editing enzymes. In addition to targeting DNA, CRISPR-Cas-based RNA-targeting tools are being developed for research, medicine and diagnostics. Nuclease-inactive and RNA-targeting Cas proteins have been fused to a plethora of effector proteins to regulate gene expression, epigenetic modifications and chromatin interactions. Collectively, the new advances are considerably improving our understanding of biological processes and are propelling CRISPR-Cas-based tools towards clinical use in gene and cell therapies.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
