

BMAL1-Driven Tissue Clocks Respond Independently to Light to Maintain Homeostasis
- PMID: 31150620
- PMCID: PMC8157517
- DOI: 10.1016/j.cell.2019.05.009
BMAL1-Driven Tissue Clocks Respond Independently to Light to Maintain Homeostasis
Erratum in
-
BMAL1-Driven Tissue Clocks Respond Independently to Light to Maintain Homeostasis.Cell. 2019 Aug 8;178(4):1029. doi: 10.1016/j.cell.2019.07.030. Cell. 2019. PMID: 31398328 Free PMC article. No abstract available.
Abstract
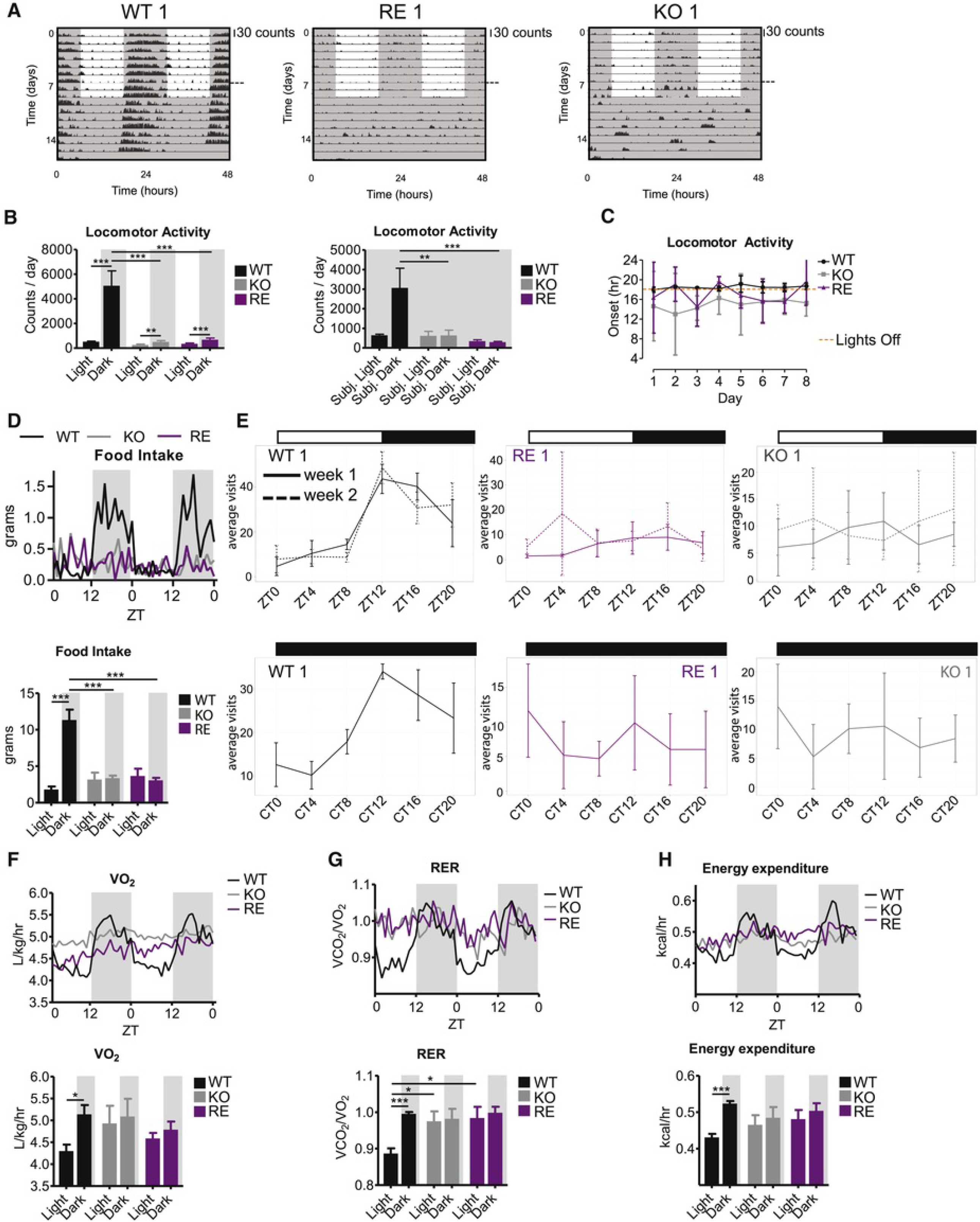
Circadian rhythms control organismal physiology throughout the day. At the cellular level, clock regulation is established by a self-sustained Bmal1-dependent transcriptional oscillator network. However, it is still unclear how different tissues achieve a synchronized rhythmic physiology. That is, do they respond independently to environmental signals, or require interactions with each other to do so? We show that unexpectedly, light synchronizes the Bmal1-dependent circadian machinery in single tissues in the absence of Bmal1 in all other tissues. Strikingly, light-driven tissue autonomous clocks occur without rhythmic feeding behavior and are lost in constant darkness. Importantly, tissue-autonomous Bmal1 partially sustains homeostasis in otherwise arrhythmic and prematurely aging animals. Our results therefore support a two-branched model for the daily synchronization of tissues: an autonomous response branch, whereby light entrains circadian clocks without any commitment of other Bmal1-dependent clocks, and a memory branch using other Bmal1-dependent clocks to "remember" time in the absence of external cues.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
