

Defining the Independence of the Liver Circadian Clock
- PMID: 31150621
- PMCID: PMC6813833
- DOI: 10.1016/j.cell.2019.04.025
Defining the Independence of the Liver Circadian Clock
Abstract
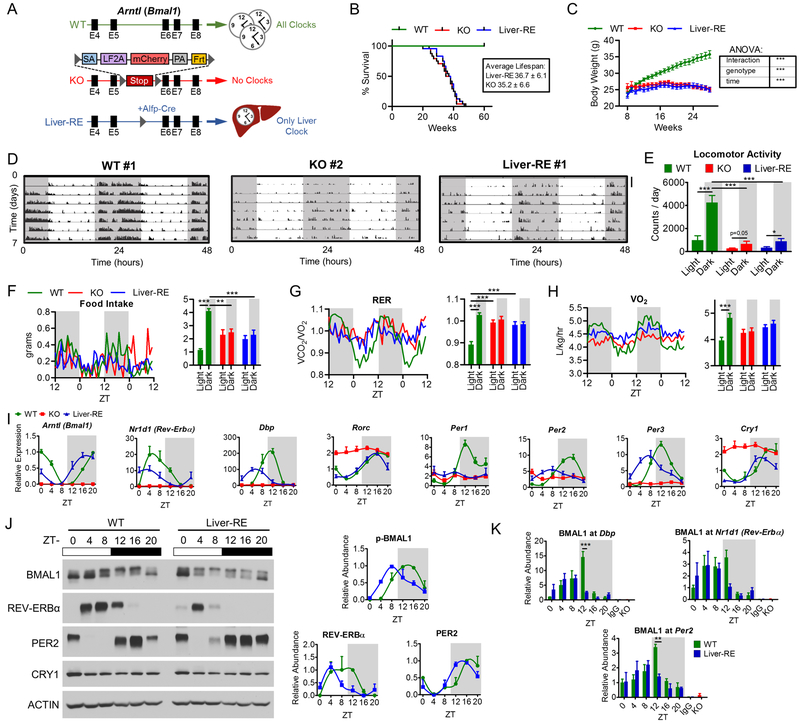
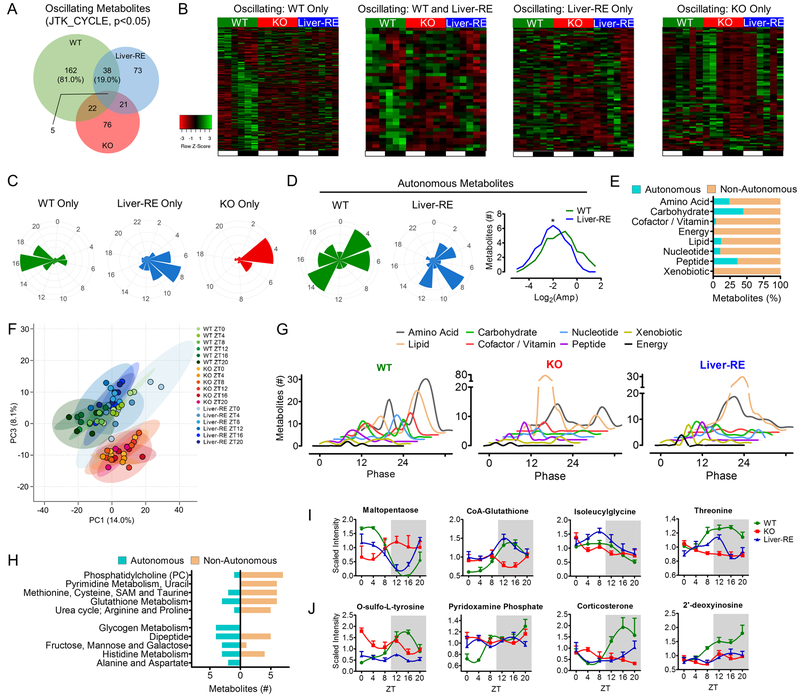
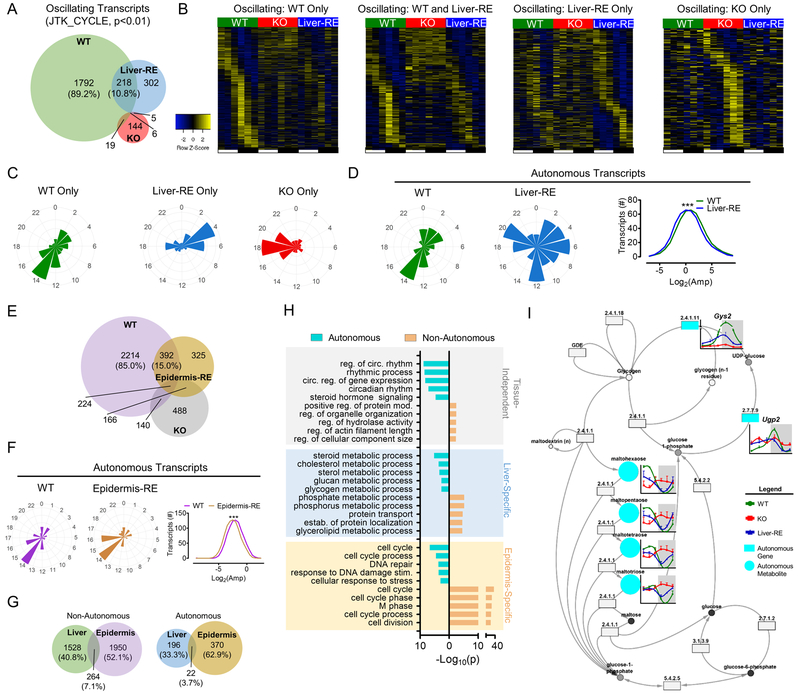
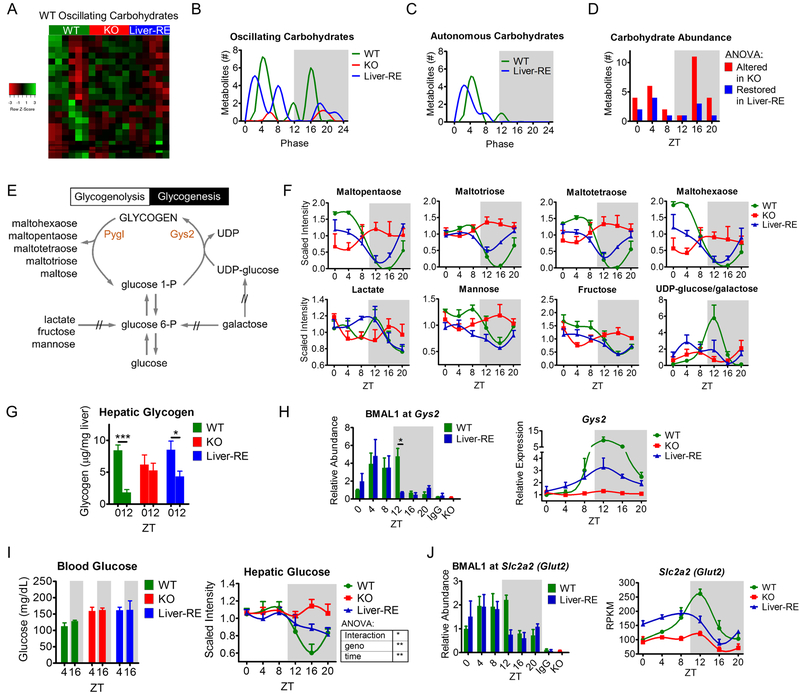
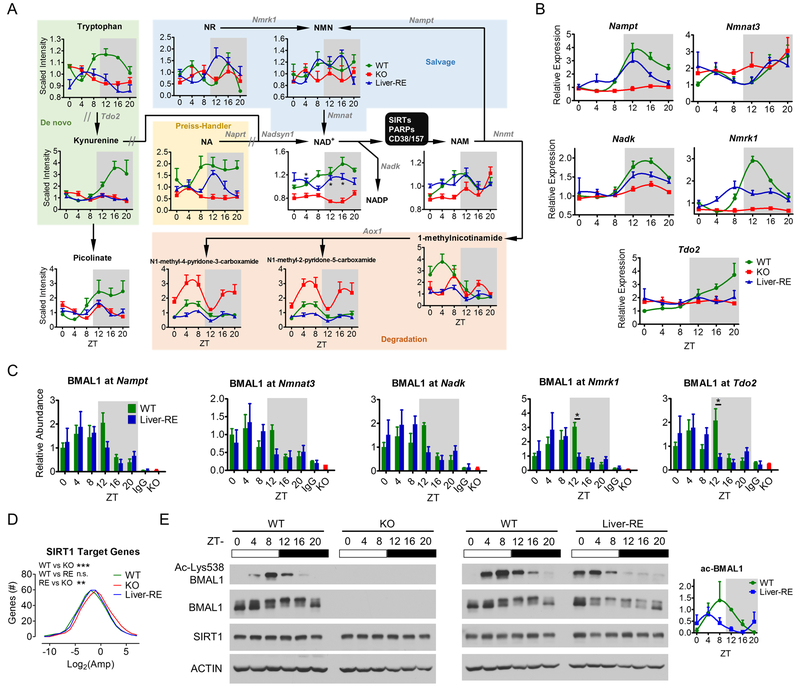
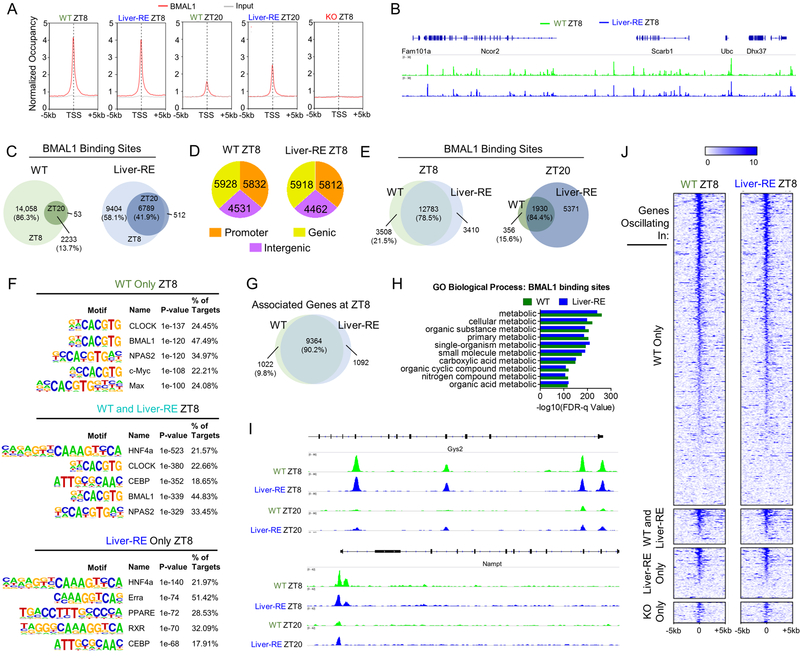
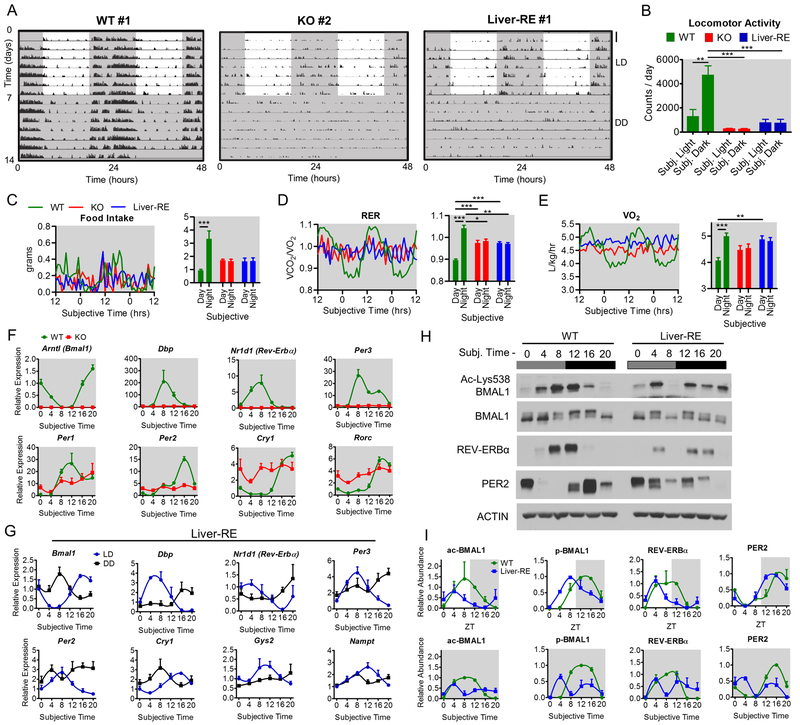
Mammals rely on a network of circadian clocks to control daily systemic metabolism and physiology. The central pacemaker in the suprachiasmatic nucleus (SCN) is considered hierarchically dominant over peripheral clocks, whose degree of independence, or tissue-level autonomy, has never been ascertained in vivo. Using arrhythmic Bmal1-null mice, we generated animals with reconstituted circadian expression of BMAL1 exclusively in the liver (Liver-RE). High-throughput transcriptomics and metabolomics show that the liver has independent circadian functions specific for metabolic processes such as the NAD+ salvage pathway and glycogen turnover. However, although BMAL1 occupies chromatin at most genomic targets in Liver-RE mice, circadian expression is restricted to ∼10% of normally rhythmic transcripts. Finally, rhythmic clock gene expression is lost in Liver-RE mice under constant darkness. Hence, full circadian function in the liver depends on signals emanating from other clocks, and light contributes to tissue-autonomous clock function.
Keywords: autonomous; bmal1; chromatin; circadian; clock; diurnal physiology; epigenetics; light; metabolism; systemic signaling.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
J.M.K is an employee of Metabolon, Inc. (no role in the study design)
Figures
Comment in
-
Reconstitution of Organismal Liver Clock Function Requires Light.Trends Endocrinol Metab. 2019 Sep;30(9):569-571. doi: 10.1016/j.tem.2019.07.017. Epub 2019 Jul 31. Trends Endocrinol Metab. 2019. PMID: 31375394
References
-
- Akhtar RA, Reddy AB, Maywood ES, Clayton JD, King VM, Smith AG, Gant TW, Hastings MH, and Kyriacou CP (2002). Circadian cycling of the mouse liver transcriptome, as revealed by cDNA microarray, is driven by the suprachiasmatic nucleus. Curr Biol 12, 540–550. - PubMed
-
- Asher G, Gatfield D, Stratmann M, Reinke H, Dibner C, Kreppel F, Mostoslavsky R, Alt FW, and Schibler U (2008). SIRT1 regulates circadian clock gene expression through PER2 deacetylation. Cell 134, 317–328. - PubMed
-
- Asher G, and Sassone-Corsi P (2015). Time for food: the intimate interplay between nutrition, metabolism, and the circadian clock. Cell 161, 84–92. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
