

A Public BCR Present in a Unique Dual-Receptor-Expressing Lymphocyte from Type 1 Diabetes Patients Encodes a Potent T Cell Autoantigen
- PMID: 31150624
- PMCID: PMC7962621
- DOI: 10.1016/j.cell.2019.05.007
A Public BCR Present in a Unique Dual-Receptor-Expressing Lymphocyte from Type 1 Diabetes Patients Encodes a Potent T Cell Autoantigen
Abstract
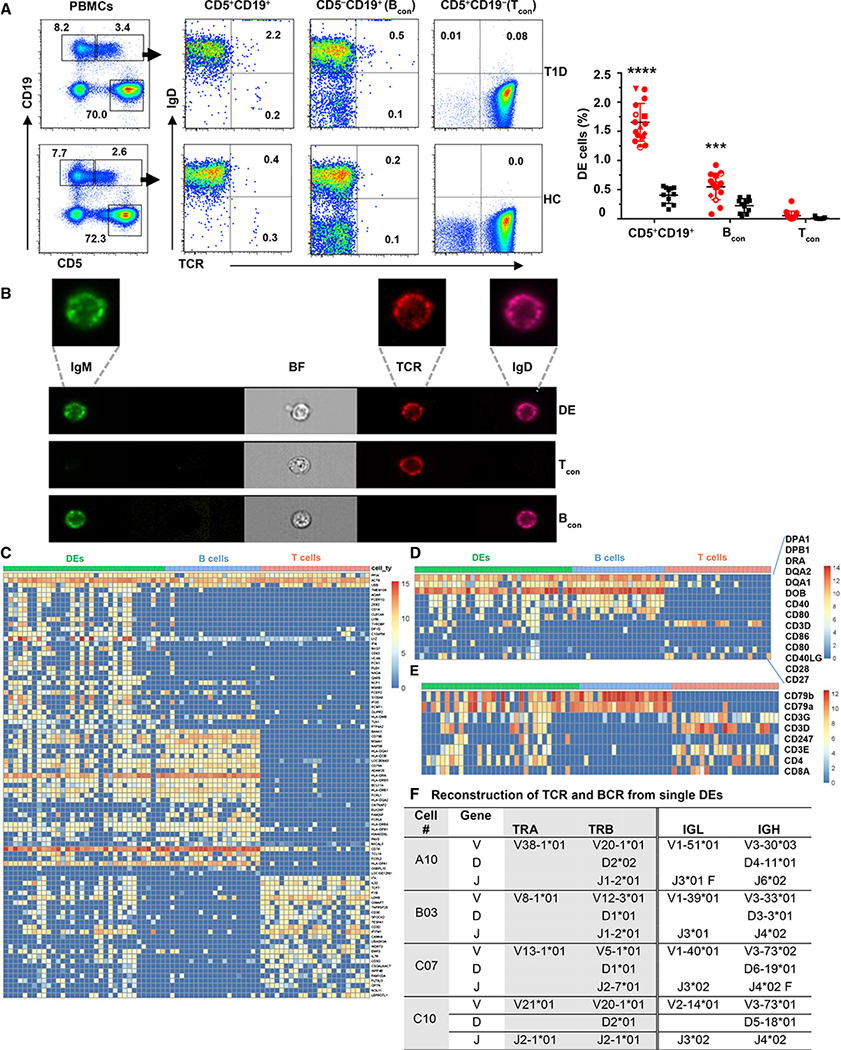
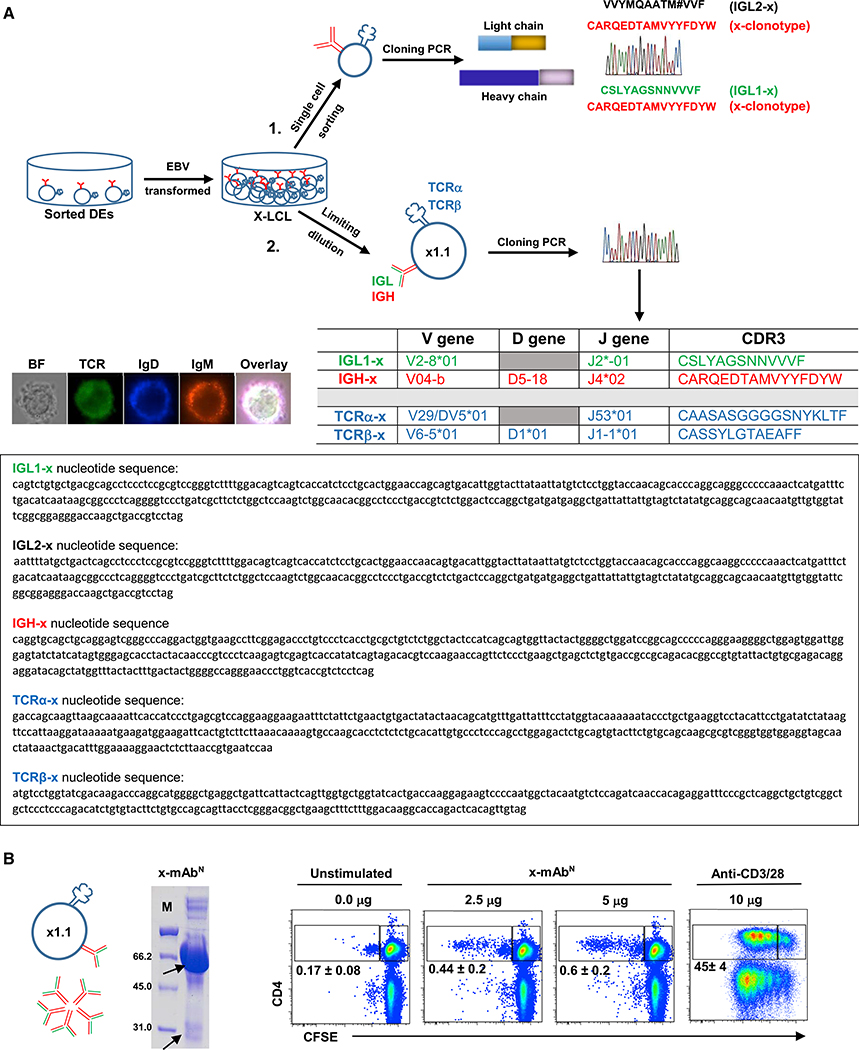
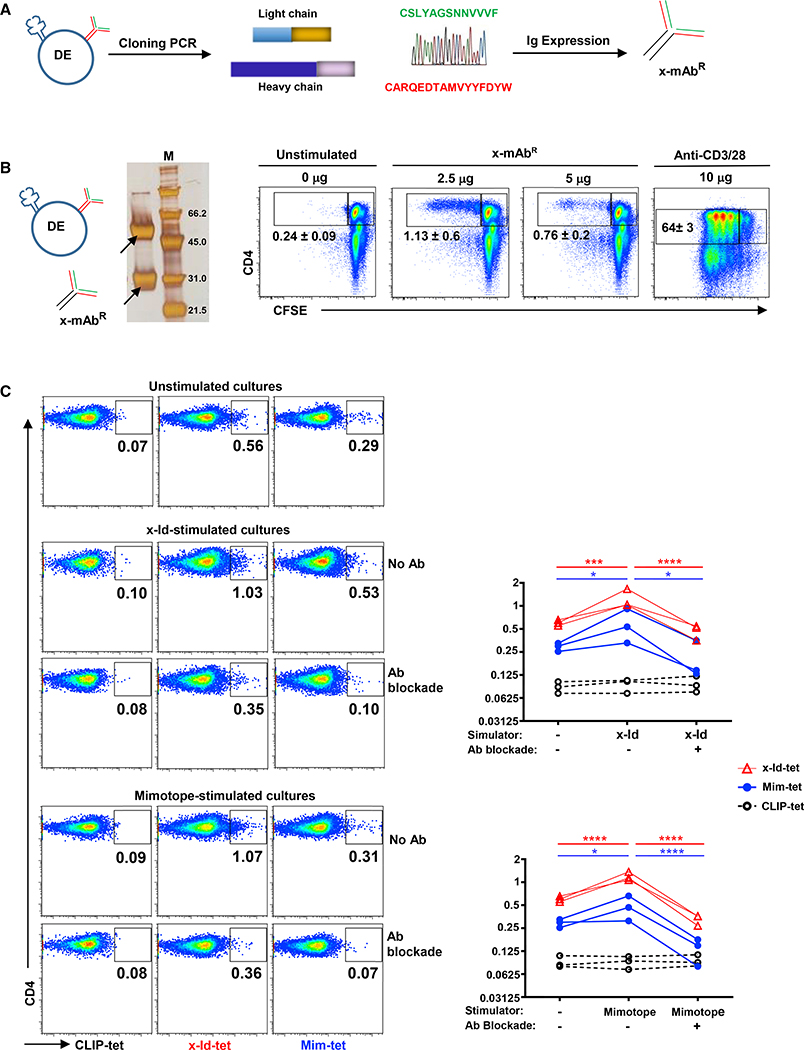
T and B cells are the two known lineages of adaptive immune cells. Here, we describe a previously unknown lymphocyte that is a dual expresser (DE) of TCR and BCR and key lineage markers of both B and T cells. In type 1 diabetes (T1D), DEs are predominated by one clonotype that encodes a potent CD4 T cell autoantigen in its antigen binding site. Molecular dynamics simulations revealed that this peptide has an optimal binding register for diabetogenic HLA-DQ8. In concordance, a synthetic version of the peptide forms stable DQ8 complexes and potently stimulates autoreactive CD4 T cells from T1D patients, but not healthy controls. Moreover, mAbs bearing this clonotype are autoreactive against CD4 T cells and inhibit insulin tetramer binding to CD4 T cells. Thus, compartmentalization of adaptive immune cells into T and B cells is not absolute, and violators of this paradigm are likely key drivers of autoimmune diseases.
Keywords: DEs; autoantigen; dual expressers; insulin mimotope; islet autoantigen; type 1 diabetes; x-clonotype; x-idiotype.
Copyright © 2019. Published by Elsevier Inc.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no conflict of interests.
Figures




Comment in
-
TCR+/BCR+ dual-expressing cells and their associated public BCR clonotype are not enriched in type 1 diabetes.Cell. 2021 Feb 4;184(3):827-839.e14. doi: 10.1016/j.cell.2020.11.035. Cell. 2021. PMID: 33545036 Free PMC article.
-
A reply to "TCR+/BCR+ dual-expressing cells and their associated public BCR clonotype are not enriched in type 1 diabetes".Cell. 2021 Feb 4;184(3):840-843. doi: 10.1016/j.cell.2020.11.036. Cell. 2021. PMID: 33545037 Free PMC article.
References
-
- Aas-Hanssen K, Funderud A, Thompson KM, Bogen B, and Munthe LA (2014). Idiotype-specific Th cells support oligoclonal expansion of anti-dsDNA B cells in mice with lupus. J. Immunol. 193, 2691–2698. - PubMed
-
- Caputo JG, and Flytzanis N (1991). Kink-antikink collisions in sine-Gordon and phi4 models: Problems in the variational approach. Phys. Rev. A 44, 6219–6225. - PubMed
-
- Carlson CS, Emerson RO, Sherwood AM, Desmarais C, Chung MW, Parsons JM, Steen MS, LaMadrid-Herrmannsfeldt MA, Williamson DW, Livingston RJ, et al. (2013). Using synthetic templates to design an unbiased multiplex PCR assay. Nat. Commun. 4, 2680. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
