

Reverse Engineering Provides Insights on the Evolution of Subgroups A to E Avian Sarcoma and Leukosis Virus Receptor Specificity
- PMID: 31151254
- PMCID: PMC6630264
- DOI: 10.3390/v11060497
Reverse Engineering Provides Insights on the Evolution of Subgroups A to E Avian Sarcoma and Leukosis Virus Receptor Specificity
Abstract
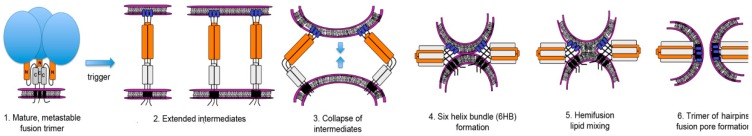
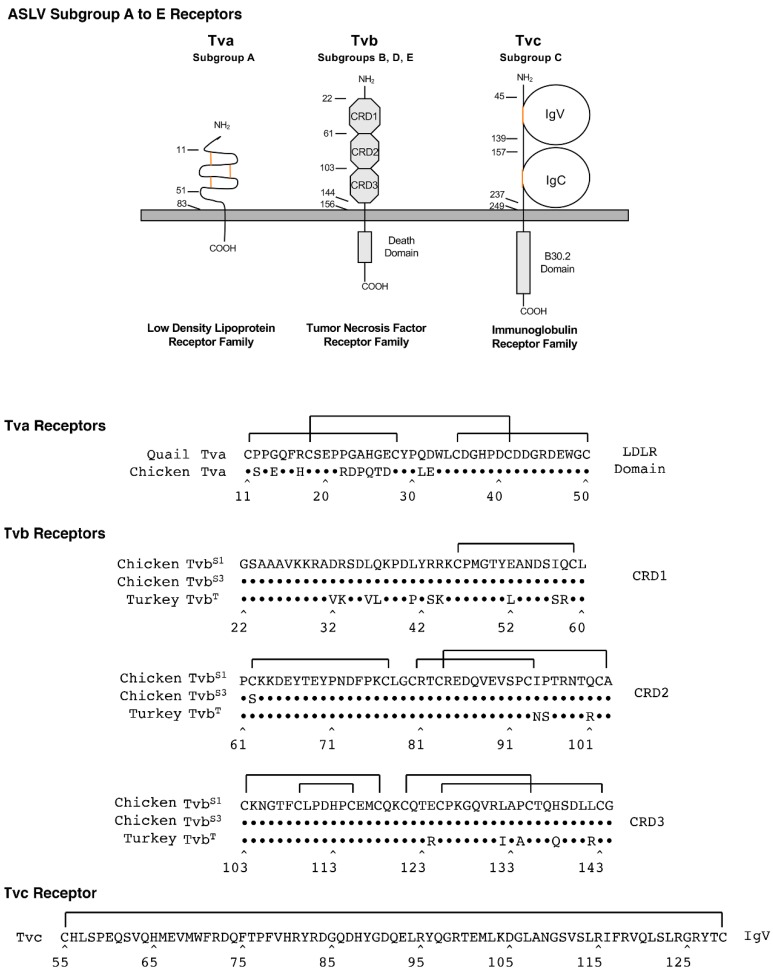
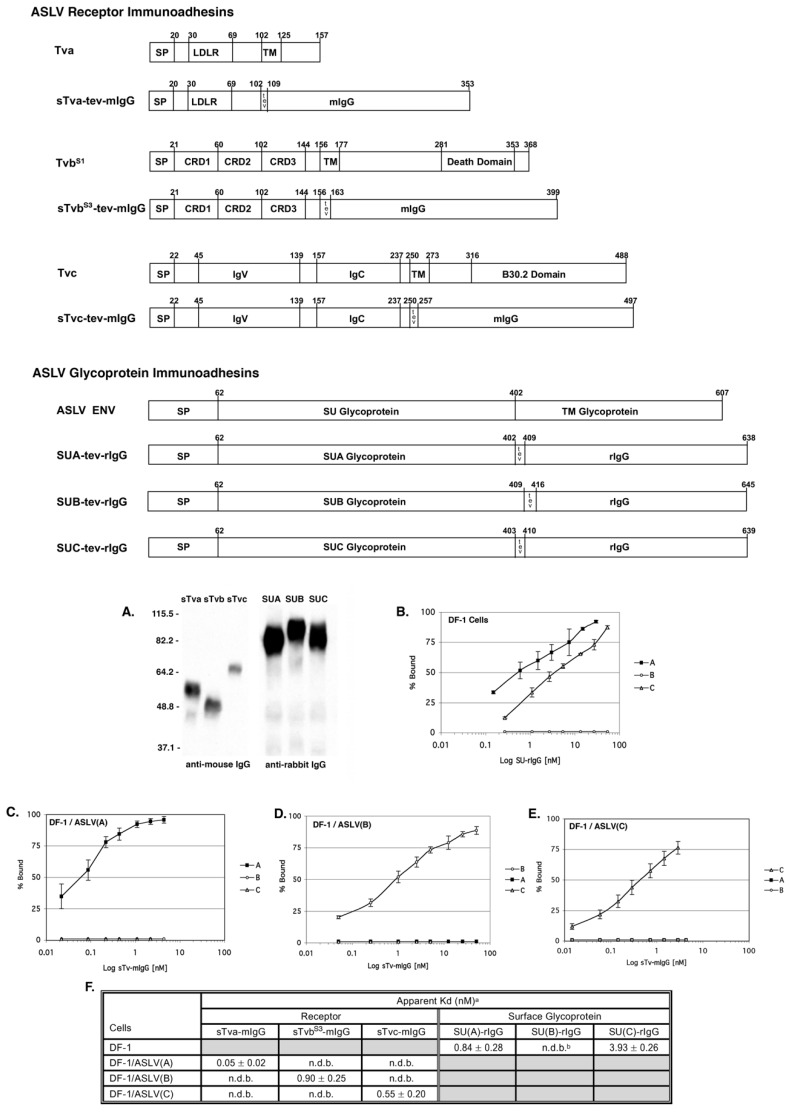
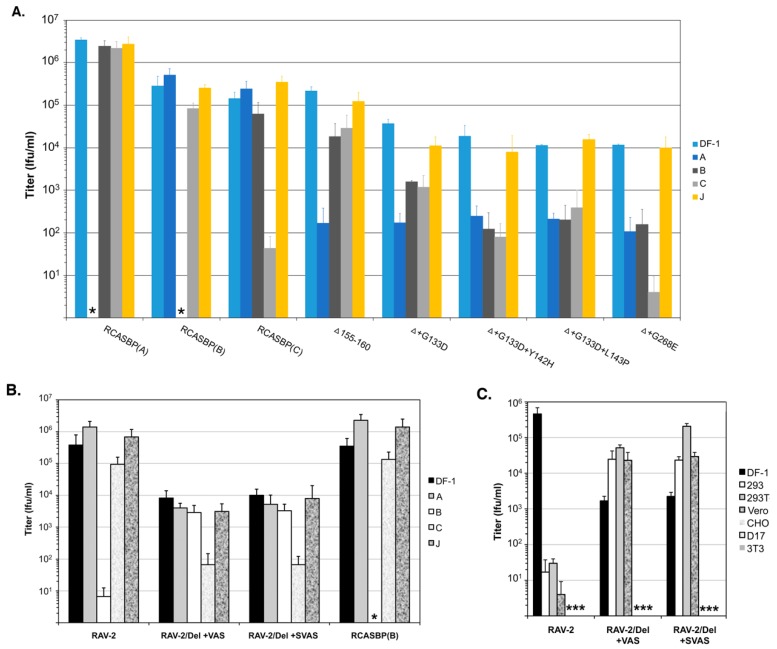
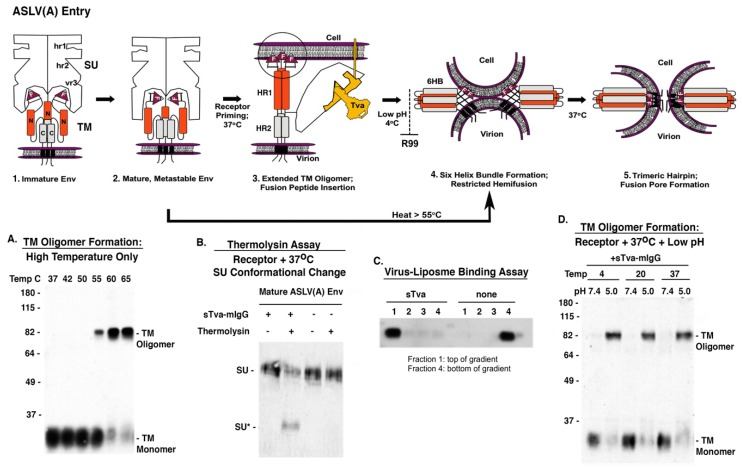
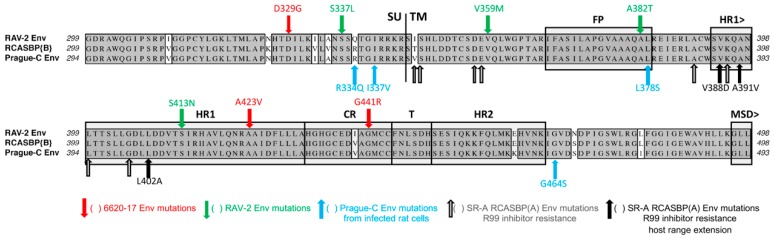
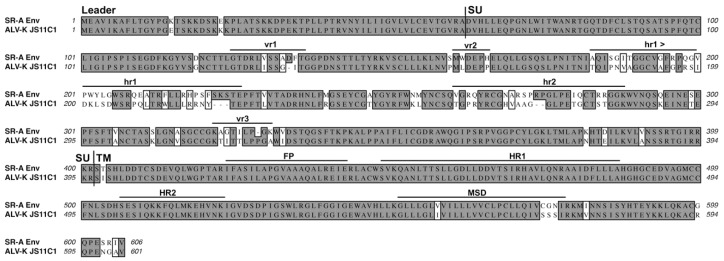
The initial step of retrovirus entry-the interaction between the virus envelope glycoprotein trimer and a cellular receptor-is complex, involving multiple, noncontiguous determinants in both proteins that specify receptor choice, binding affinity and the ability to trigger conformational changes in the viral glycoproteins. Despite the complexity of this interaction, retroviruses have the ability to evolve the structure of their envelope glycoproteins to use a different cellular protein as receptors. The highly homologous subgroup A to E Avian Sarcoma and Leukosis Virus (ASLV) glycoproteins belong to the group of class 1 viral fusion proteins with a two-step triggering mechanism that allows experimental access to intermediate structures during the fusion process. We and others have taken advantage of replication-competent ASLVs and exploited genetic selection strategies to force the ASLVs to naturally evolve and acquire envelope glycoprotein mutations to escape the pressure on virus entry and still yield a functional replicating virus. This approach allows for the simultaneous selection of multiple mutations in multiple functional domains of the envelope glycoprotein that may be required to yield a functional virus. Here, we review the ASLV family and experimental system and the reverse engineering approaches used to understand the evolution of ASLV receptor usage.
Keywords: Avian Sarcoma and Leukosis Viruses; envelope glycoprotein evolution; receptor usage.
Conflict of interest statement
The author declares no conflicts of interest.
Figures



Similar articles
-
Mutations in Both the Surface and Transmembrane Envelope Glycoproteins of the RAV-2 Subgroup B Avian Sarcoma and Leukosis Virus Are Required to Escape the Antiviral Effect of a Secreted Form of the TvbS3 Receptor †.Viruses. 2019 May 31;11(6):500. doi: 10.3390/v11060500. Viruses. 2019. PMID: 31159208 Free PMC article.
-
Model of the TVA receptor determinants required for efficient infection by subgroup A avian sarcoma and leukosis viruses.J Virol. 2015 Feb;89(4):2136-48. doi: 10.1128/JVI.02339-14. Epub 2014 Dec 3. J Virol. 2015. PMID: 25473063 Free PMC article.
-
Avian Sarcoma and Leukosis Virus Envelope Glycoproteins Evolve to Broaden Receptor Usage Under Pressure from Entry Competitors †.Viruses. 2019 Jun 5;11(6):519. doi: 10.3390/v11060519. Viruses. 2019. PMID: 31195660 Free PMC article.
-
Alpharetrovirus envelope-receptor interactions.Curr Top Microbiol Immunol. 2003;281:107-36. doi: 10.1007/978-3-642-19012-4_3. Curr Top Microbiol Immunol. 2003. PMID: 12932076 Review.
-
Avian sarcoma and leukosis virus-receptor interactions: from classical genetics to novel insights into virus-cell membrane fusion.Virology. 2006 Jan 5;344(1):25-9. doi: 10.1016/j.virol.2005.09.021. Virology. 2006. PMID: 16364732 Review.
Cited by
-
N123I mutation in the ALV-J receptor-binding domain region enhances viral replication ability by increasing the binding affinity with chNHE1.PLoS Pathog. 2024 Feb 7;20(2):e1011928. doi: 10.1371/journal.ppat.1011928. eCollection 2024 Feb. PLoS Pathog. 2024. PMID: 38324558 Free PMC article.
-
Novel mutation of avian leukosis virus subgroup J from Tibetan chickens.Poult Sci. 2021 Mar;100(3):100931. doi: 10.1016/j.psj.2020.12.028. Epub 2021 Jan 29. Poult Sci. 2021. PMID: 33518331 Free PMC article.
-
Simultaneous Detection of Three Subgroups of Avian Leukosis Virus Using the Nanoparticle-Assisted PCR Assay.Viruses. 2023 Dec 21;16(1):15. doi: 10.3390/v16010015. Viruses. 2023. PMID: 38275950 Free PMC article.
-
Residues L55 and W69 of Tva Mediate Entry of Subgroup A Avian Leukosis Virus.J Virol. 2022 Sep 28;96(18):e0067822. doi: 10.1128/jvi.00678-22. Epub 2022 Sep 7. J Virol. 2022. PMID: 36069550 Free PMC article.
-
Single Amino Acids G196 and R198 in hr1 of Subgroup K Avian Leukosis Virus Glycoprotein Are Critical for Tva Receptor Binding.Front Microbiol. 2020 Dec 16;11:596586. doi: 10.3389/fmicb.2020.596586. eCollection 2020. Front Microbiol. 2020. PMID: 33391214 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
