

Rapid Plasticity of Higher-Order Thalamocortical Inputs during Sensory Learning
- PMID: 31151774
- PMCID: PMC10038228
- DOI: 10.1016/j.neuron.2019.04.037
Rapid Plasticity of Higher-Order Thalamocortical Inputs during Sensory Learning
Abstract
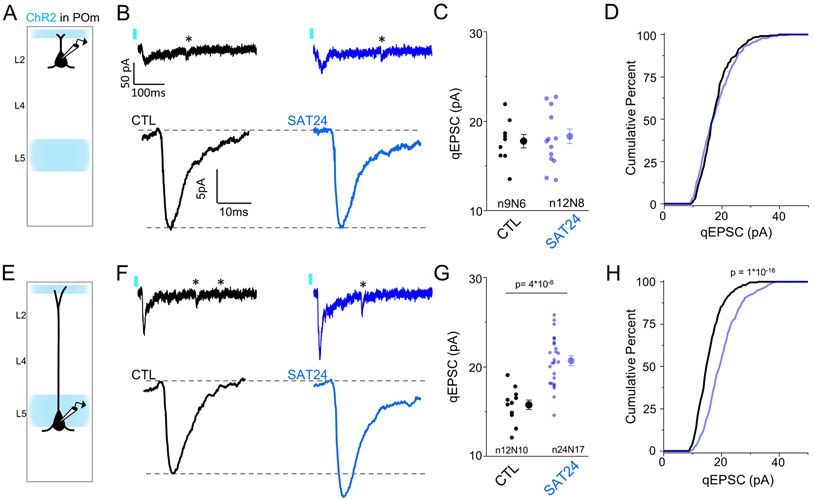
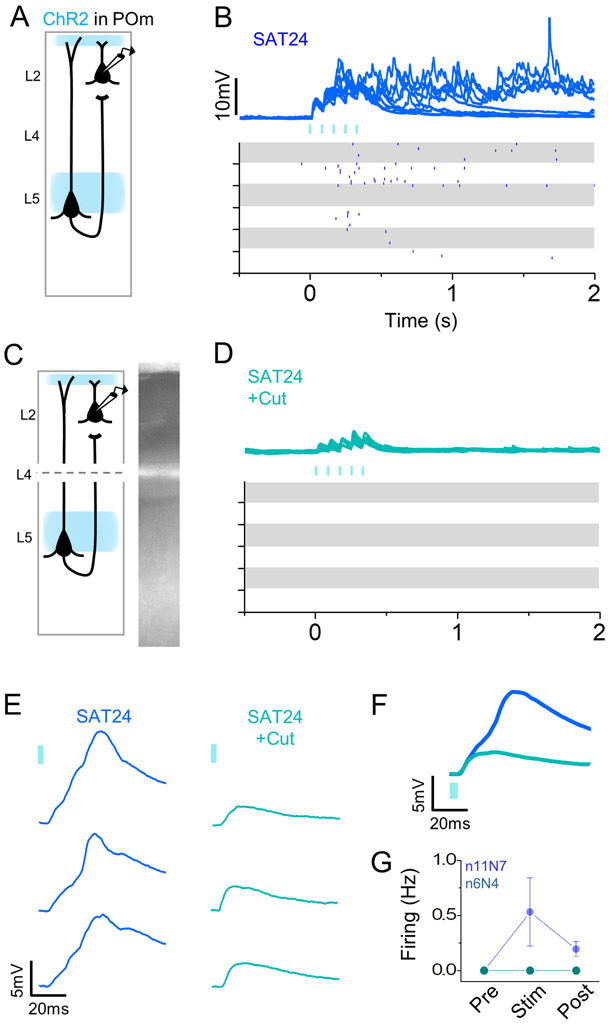
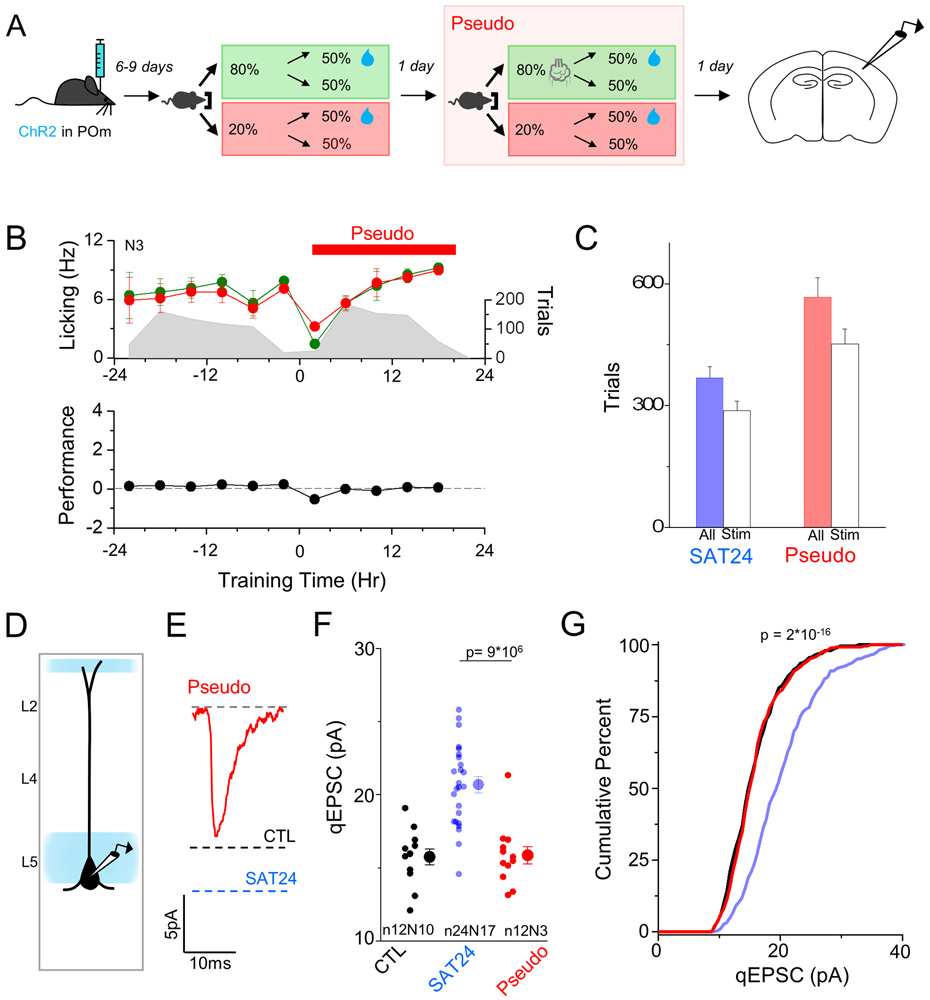
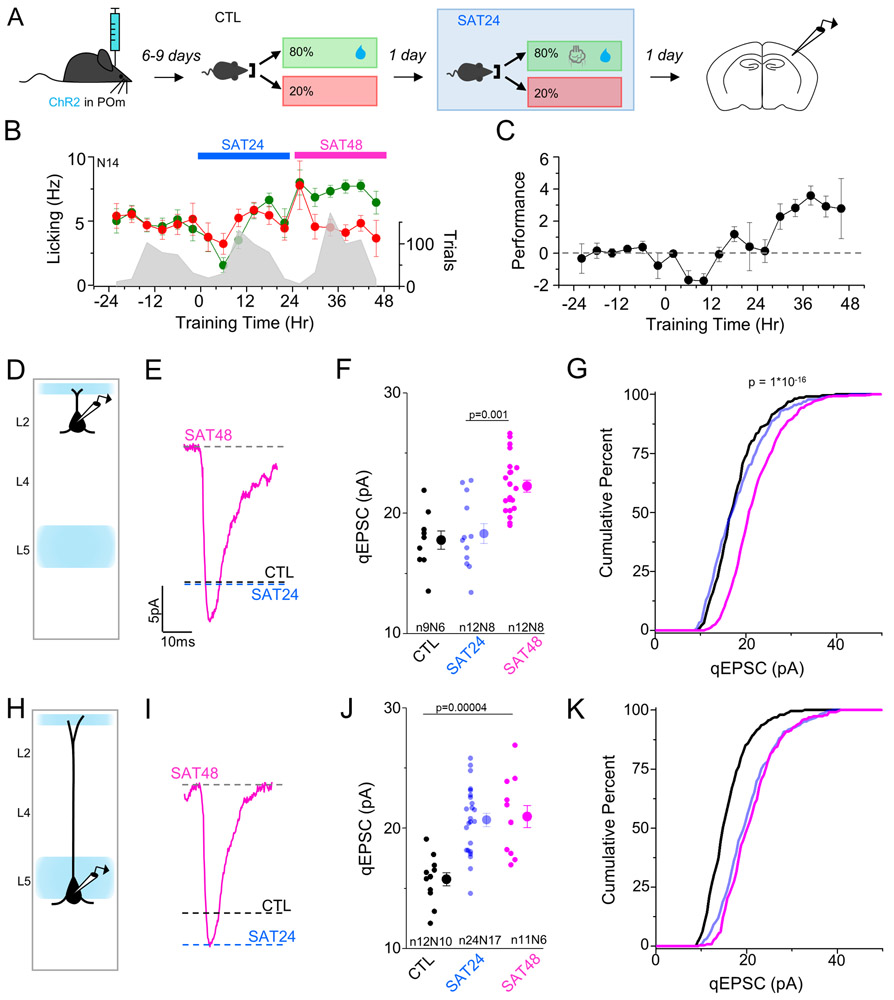
Neocortical circuits are sensitive to experience, showing both anatomical and electrophysiological changes in response to altered sensory input. We examined input- and cell-type-specific changes in thalamo- and intracortical pathways during learning using an automated, home-cage sensory association training (SAT) paradigm coupling multi-whisker stimulation to a water reward. We found that the posterior medial nucleus (POm) but not the ventral posterior medial (VPM) nucleus of the thalamus drives increased cortical activity after 24 h of SAT, when behavioral evidence of learning first emerges. Synaptic strengthening within the POm thalamocortical pathway was first observed at thalamic inputs to L5 and was not generated by sensory stimulation alone. Synaptic changes in L2 were delayed relative to L5, requiring 48 h of SAT to drive synaptic plasticity at thalamic and intracortical inputs onto L2 Pyr neurons. These data identify the POm thalamocortical circuit as a site of rapid synaptic plasticity during learning and suggest a temporal sequence to learning-evoked synaptic changes in the sensory cortex.
Copyright © 2019. Published by Elsevier Inc.
Figures
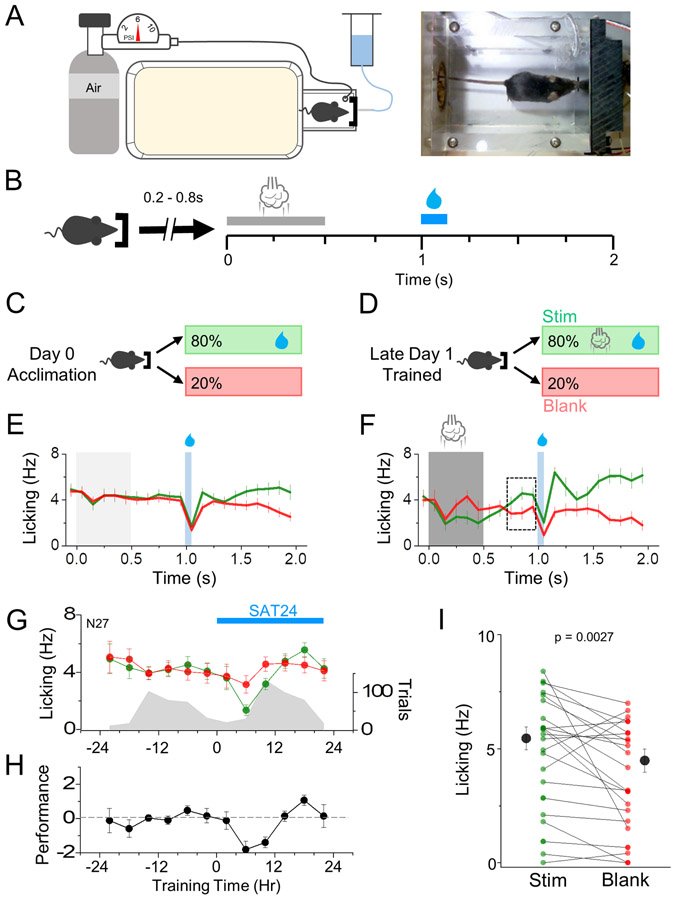
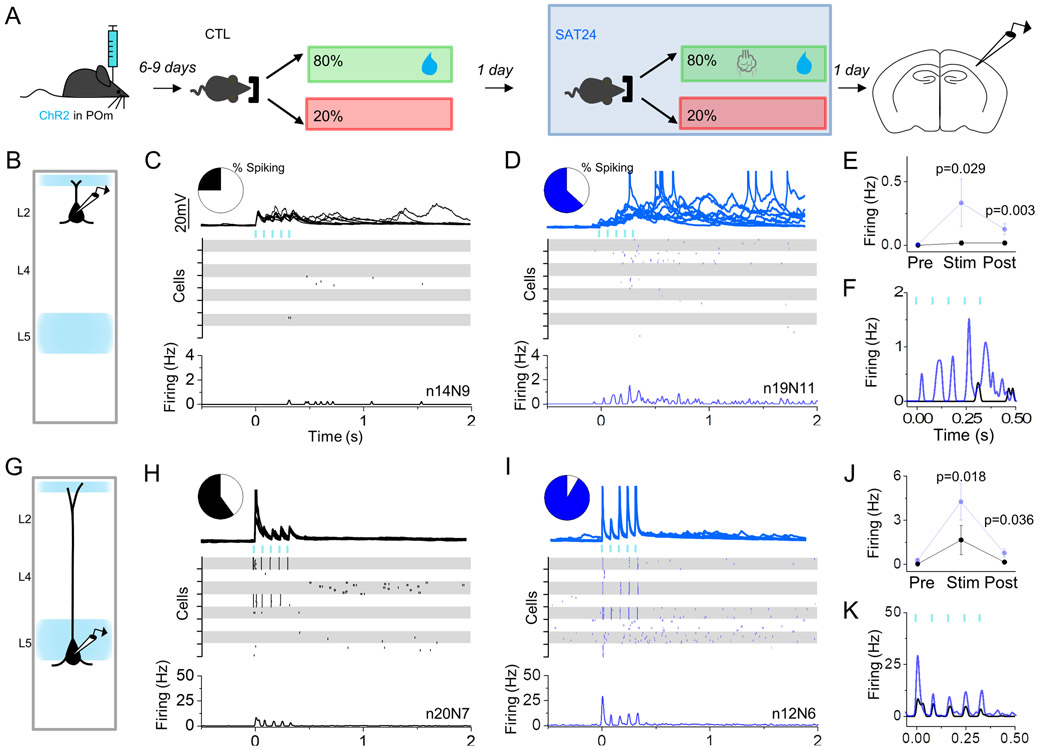
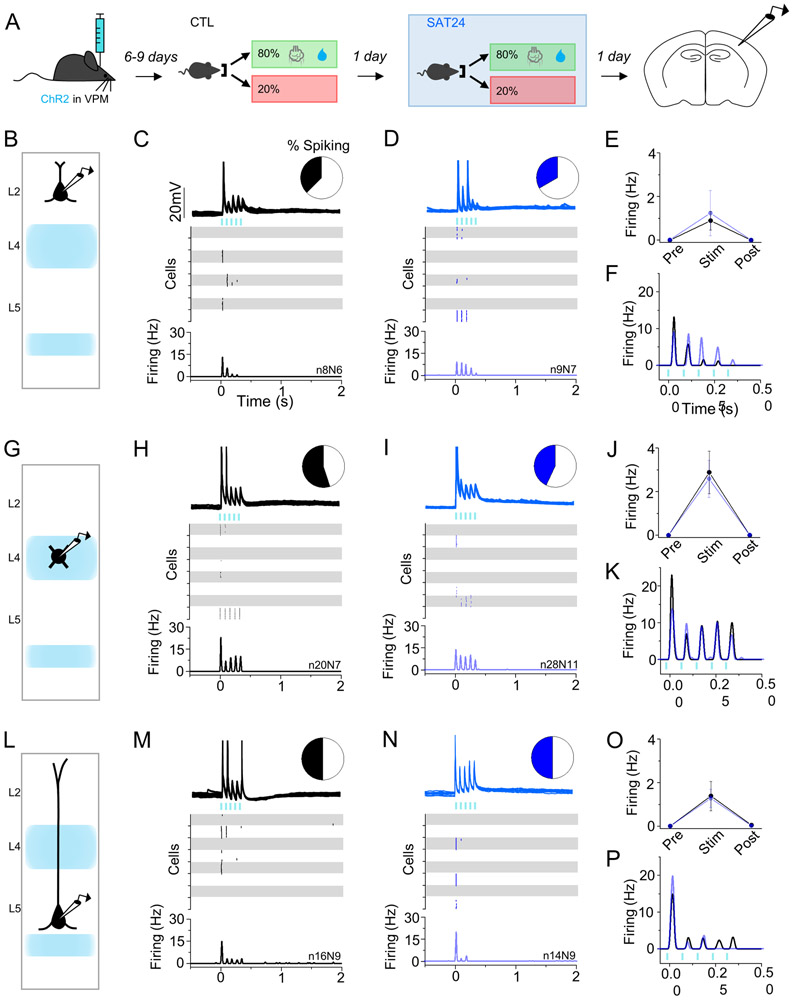
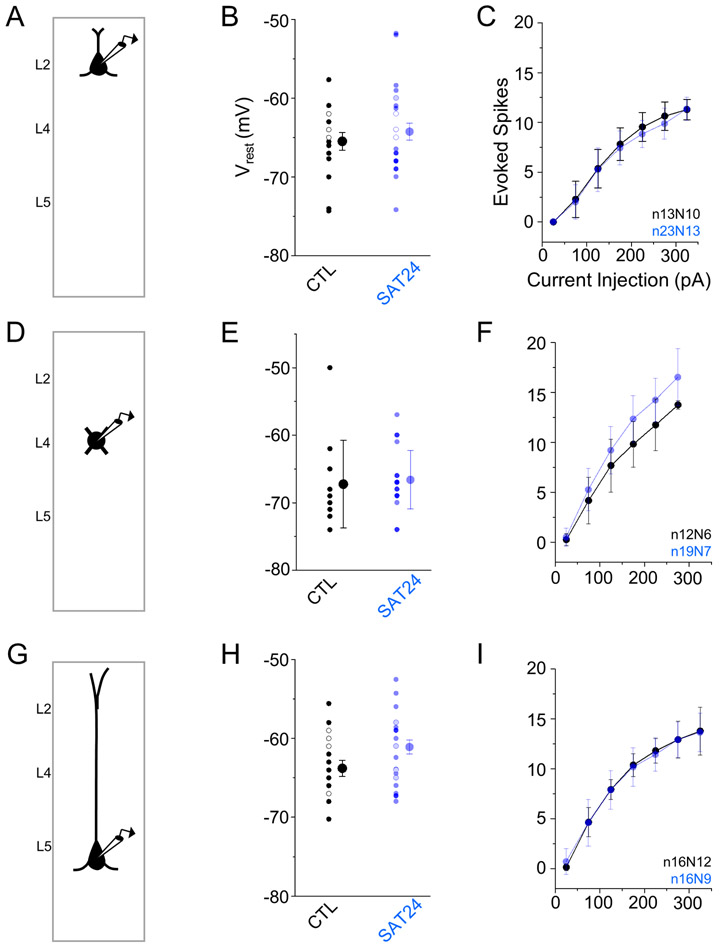
Comment in
-
Traces of Learning in Thalamocortical Circuits.Neuron. 2019 Jul 17;103(2):175-176. doi: 10.1016/j.neuron.2019.06.020. Neuron. 2019. PMID: 31319043
References
-
- Alloway KD, Hoffer ZS, and Hoover JE (2003). Quantitative comparisons of corticothalamic topography within the ventrobasal complex and the posterior nucleus of the rodent thalamus. Brain Res 968, 54–68. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
