

Arabidopsis ABCG28 is required for the apical accumulation of reactive oxygen species in growing pollen tubes
- PMID: 31152136
- PMCID: PMC6589667
- DOI: 10.1073/pnas.1902010116
Arabidopsis ABCG28 is required for the apical accumulation of reactive oxygen species in growing pollen tubes
Abstract
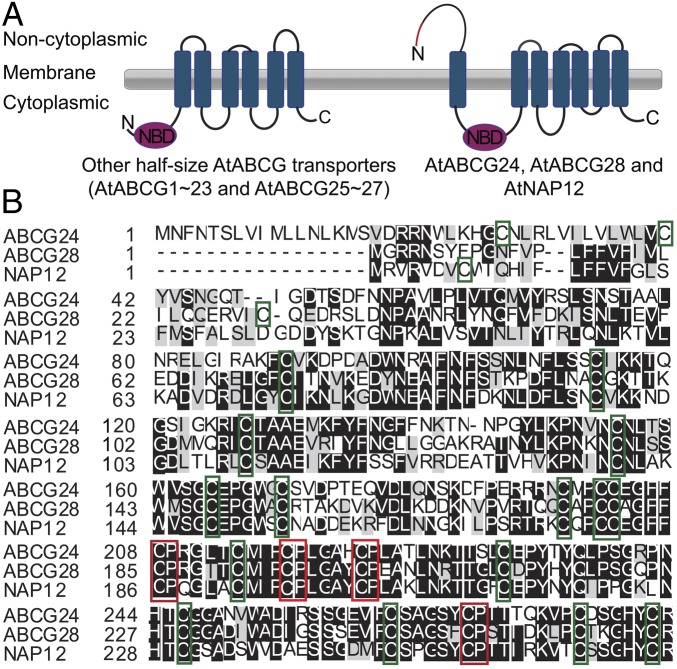
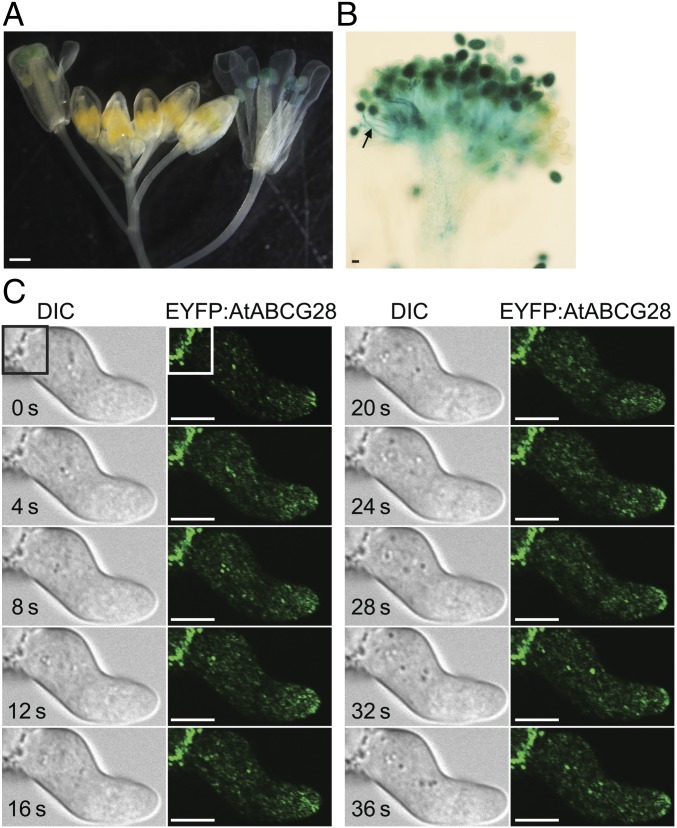
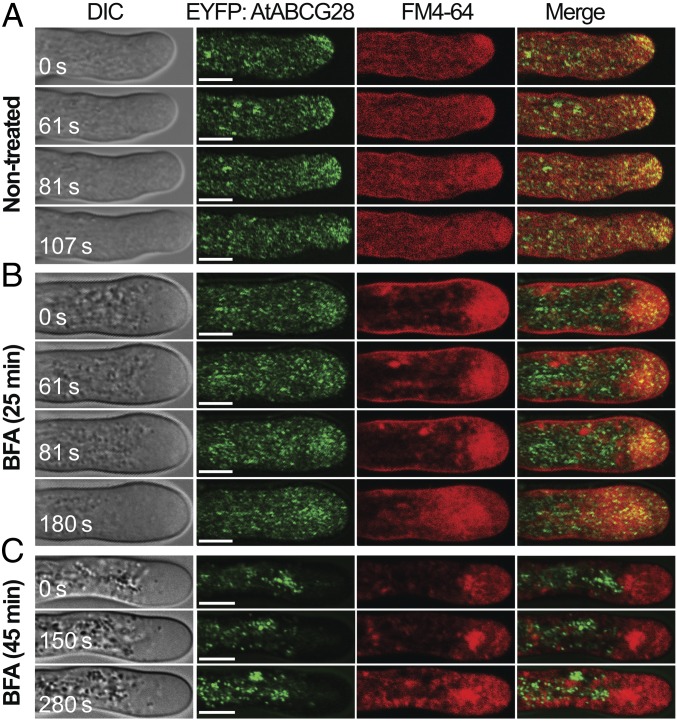
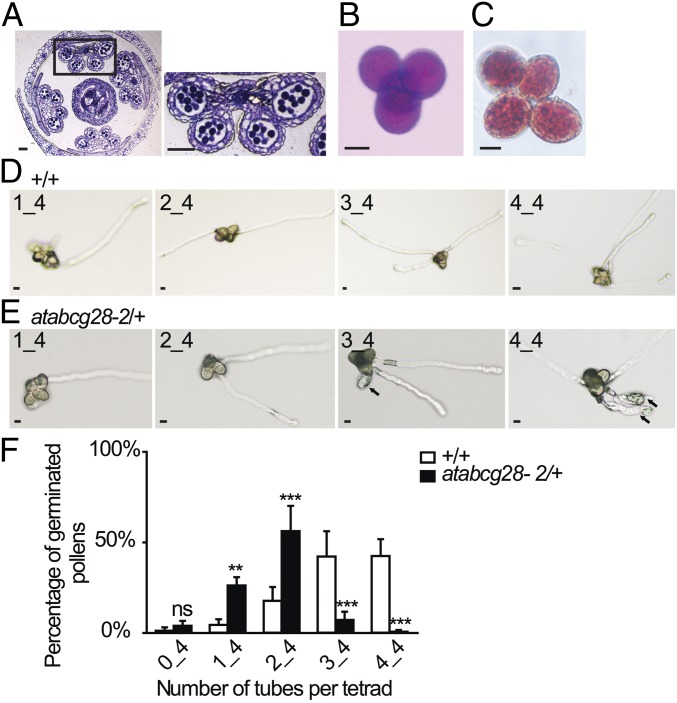
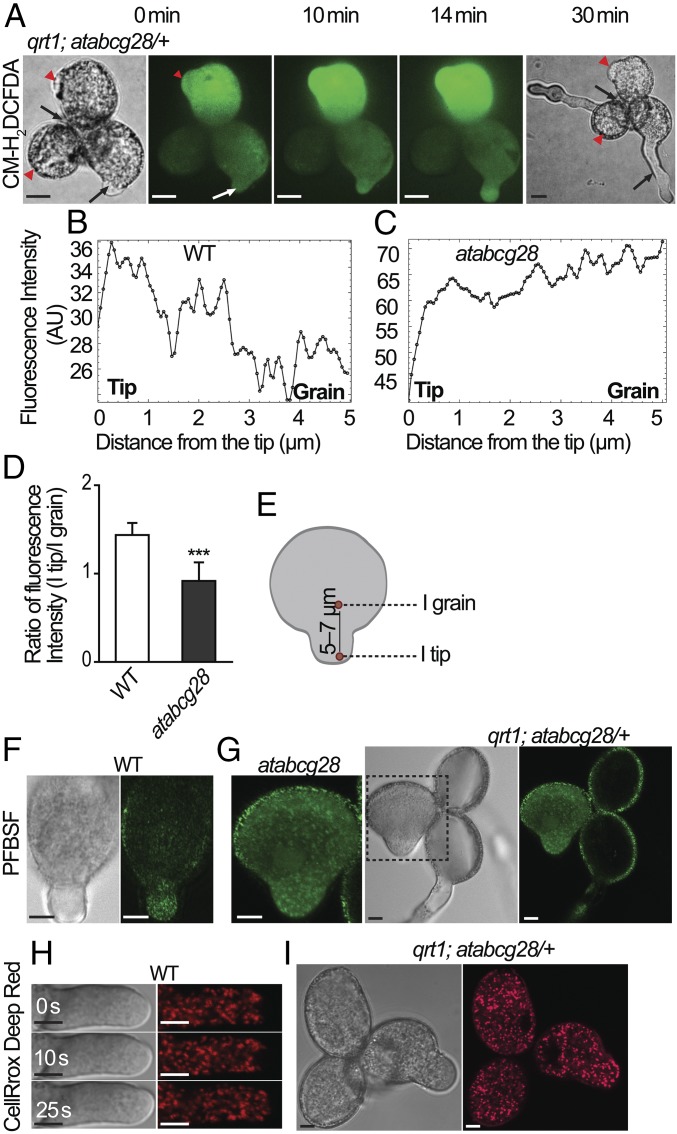
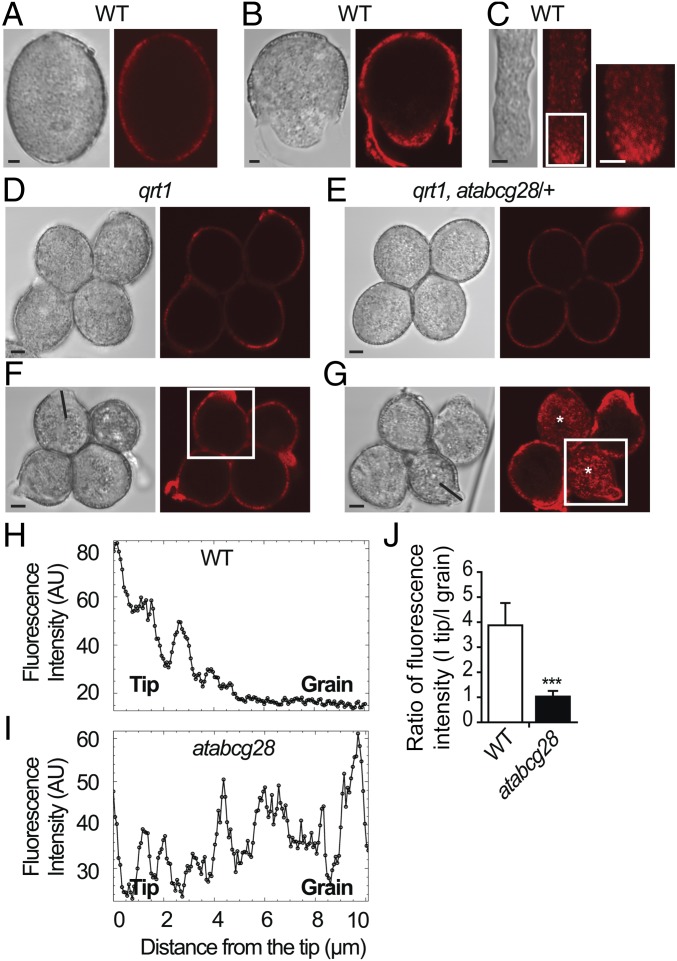
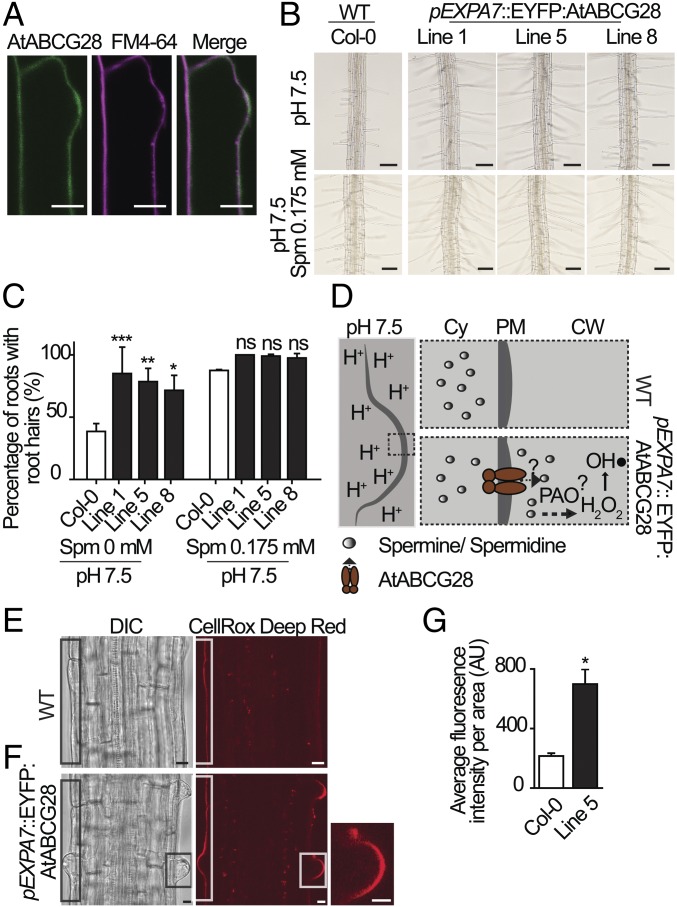
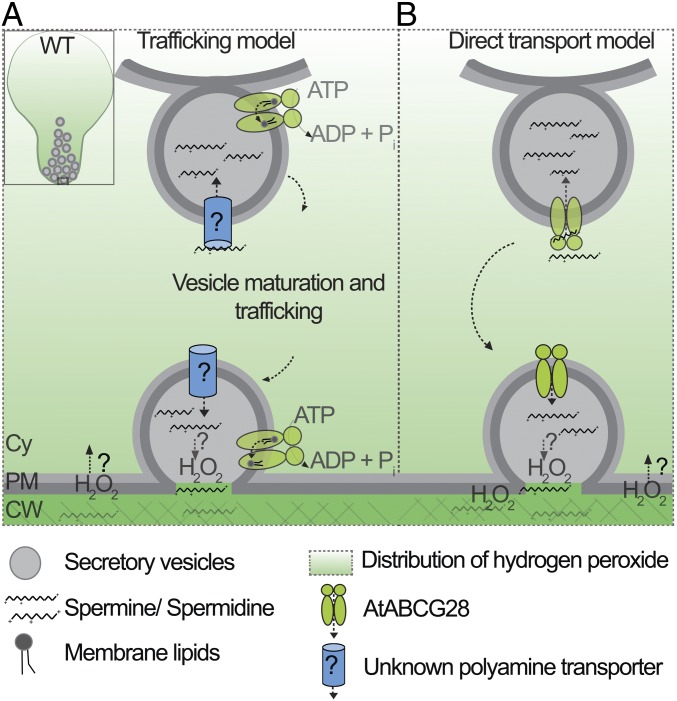
Tip-focused accumulation of reactive oxygen species (ROS) is tightly associated with pollen tube growth and is thus critical for fertilization. However, it is unclear how tip-growing cells establish such specific ROS localization. Polyamines have been proposed to function in tip growth as precursors of the ROS, hydrogen peroxide. The ABC transporter AtABCG28 may regulate ROS status, as it contains multiple cysteine residues, a characteristic of proteins involved in ROS homeostasis. In this study, we found that AtABCG28 was specifically expressed in the mature pollen grains and pollen tubes. AtABCG28 was localized to secretory vesicles inside the pollen tube that moved toward and fused with the plasma membrane of the pollen tube tip. Knocking out AtABCG28 resulted in defective pollen tube growth, failure to localize polyamine and ROS to the growing pollen tube tip, and complete male sterility, whereas ectopic expression of this gene in root hair could recover ROS accumulation at the tip and improved the growth under high-pH conditions, which normally prevent ROS accumulation and tip growth. Together, these data suggest that AtABCG28 is critical for localizing polyamine and ROS at the growing tip. In addition, this function of AtABCG28 is likely to protect the pollen tube from the cytotoxicity of polyamine and contribute to the delivery of polyamine to the growing tip for incorporation into the expanding cell wall.
Keywords: ABC transporters; AtABCG28; ROS; pollen tube growth; polyamine.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Neill S., Desikan R., Hancock J., Hydrogen peroxide signalling. Curr. Opin. Plant Biol. 5, 388–395 (2002). - PubMed
-
- Coelho S. M., Brownlee C., Bothwell J. H., A tip-high, Ca(2+) -interdependent, reactive oxygen species gradient is associated with polarized growth in fucus serratus zygotes. Planta 227, 1037–1046 (2008). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
