

The ParB homologs, Spo0J and Noc, together prevent premature midcell Z ring assembly when the early stages of replication are blocked in Bacillus subtilis
- PMID: 31152469
- PMCID: PMC6852036
- DOI: 10.1111/mmi.14319
The ParB homologs, Spo0J and Noc, together prevent premature midcell Z ring assembly when the early stages of replication are blocked in Bacillus subtilis
Abstract
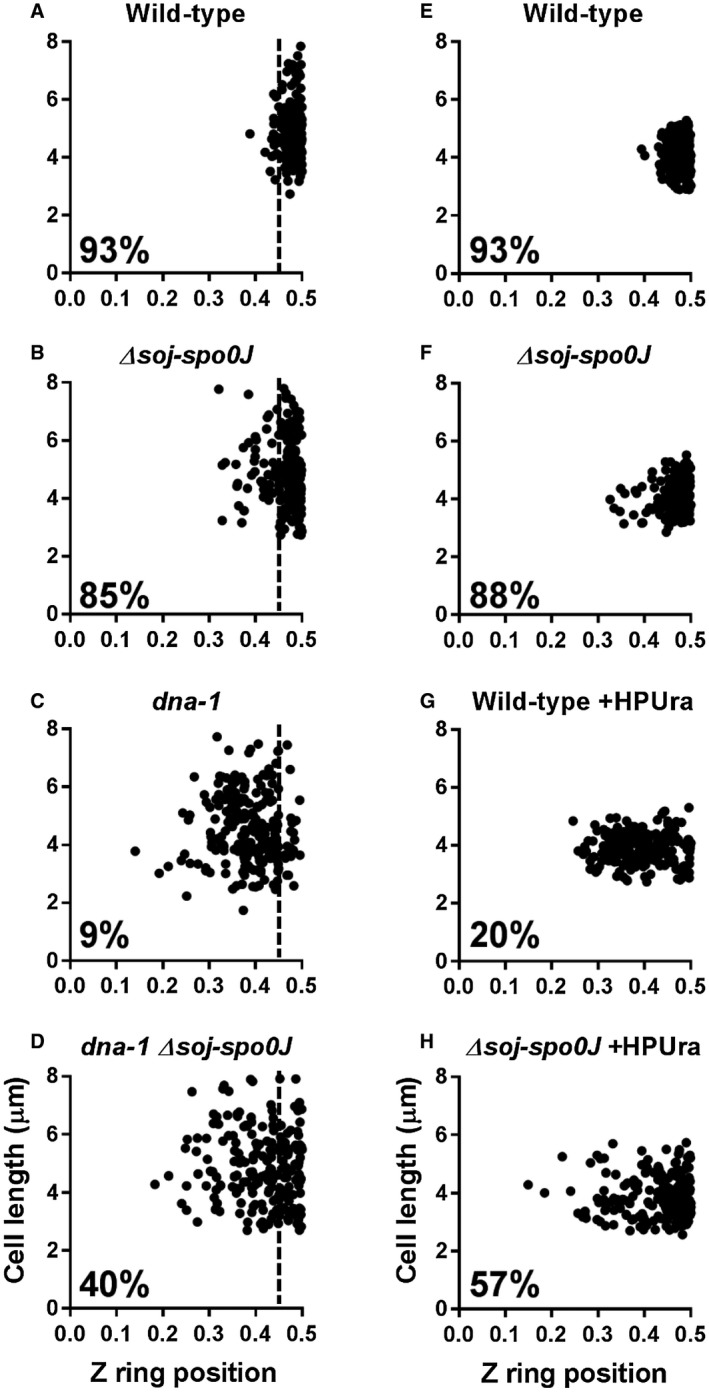
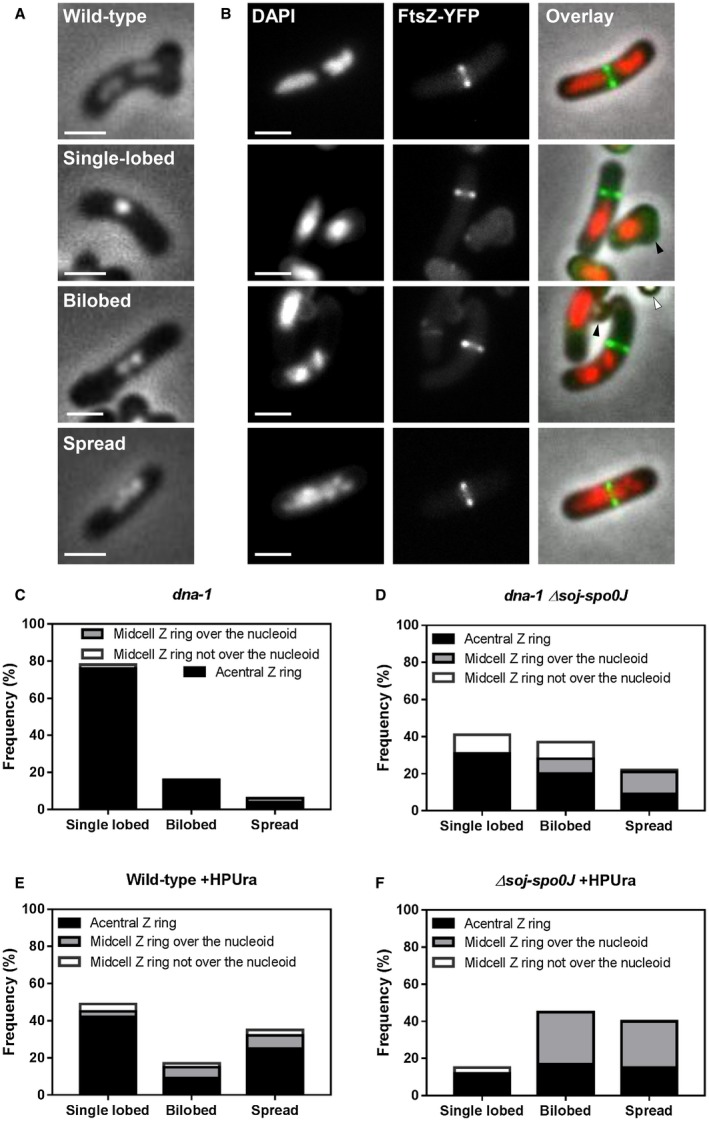
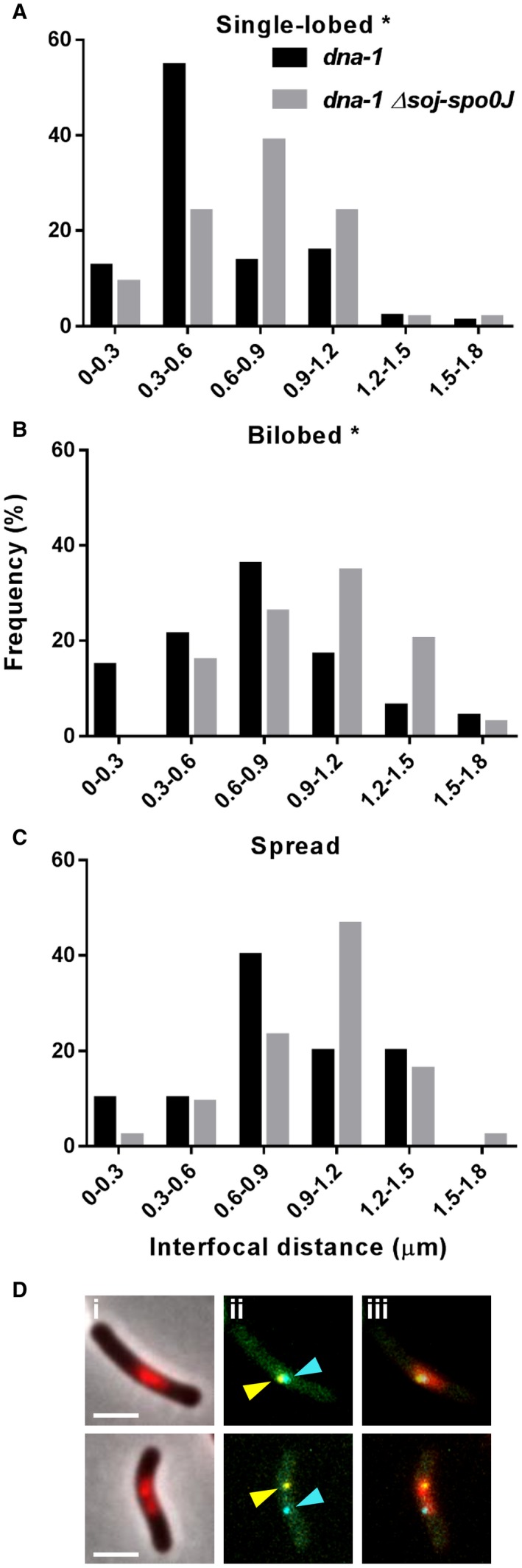
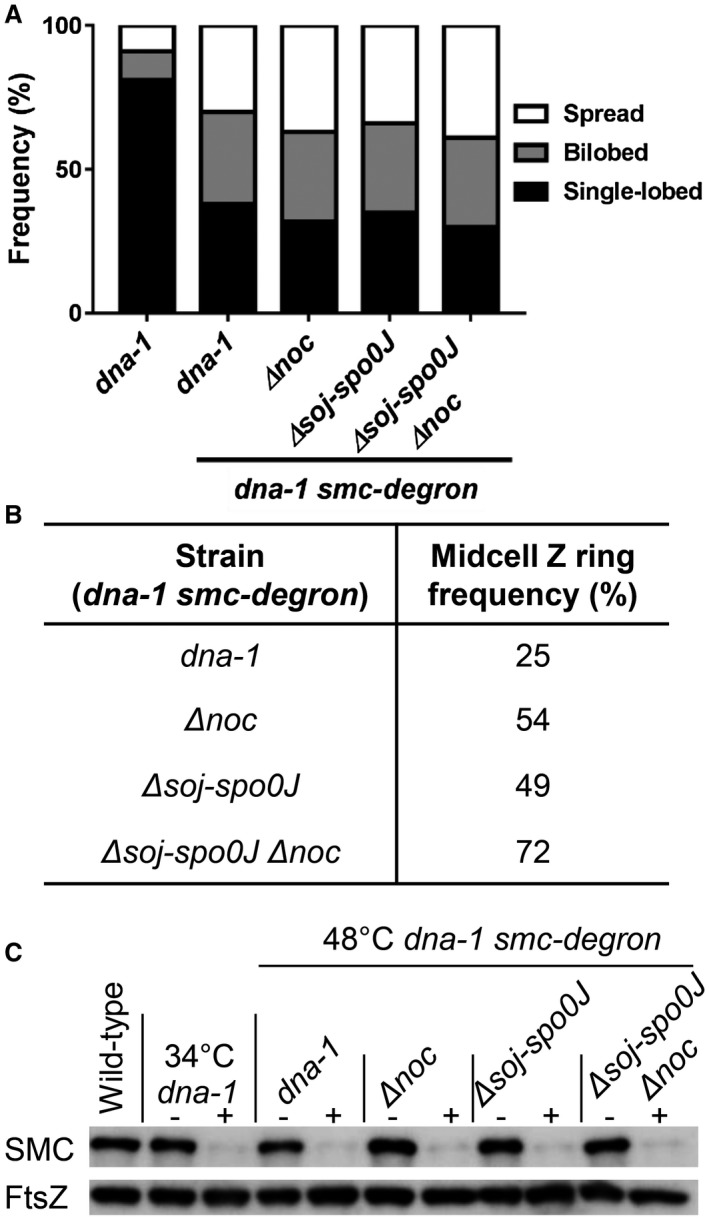
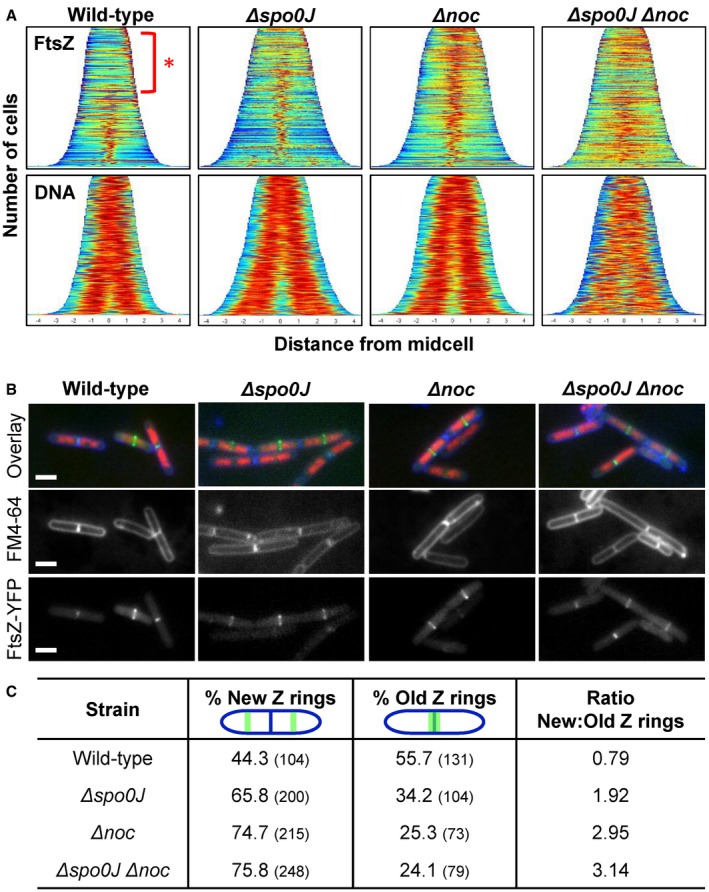
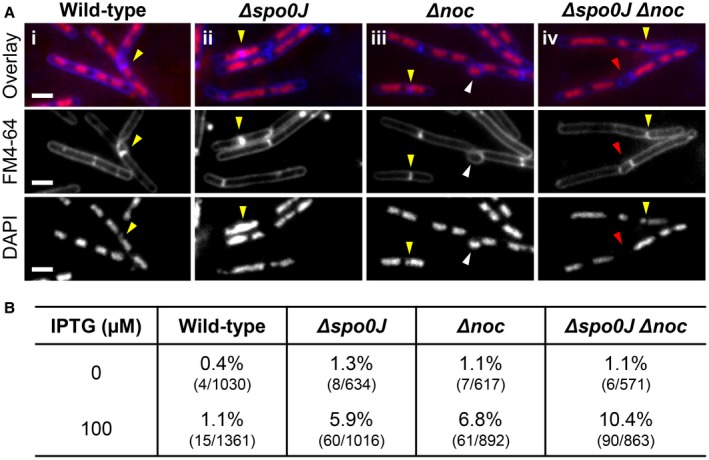
Precise cell division in coordination with DNA replication and segregation is of utmost importance for all organisms. The earliest stage of cell division is the assembly of a division protein FtsZ into a ring, known as the Z ring, at midcell. What still eludes us, however, is how bacteria precisely position the Z ring at midcell. Work in B. subtilis over the last two decades has identified a link between the early stages of DNA replication and cell division. A recent model proposed that the progression of the early stages of DNA replication leads to an increased ability for the Z ring to form at midcell. This model arose through studies examining Z ring position in mutants blocked at different steps of the early stages of DNA replication. Here, we show that this model is unlikely to be correct and the mutants previously studied generate nucleoids with different capacity for blocking midcell Z ring assembly. Importantly, our data suggest that two proteins of the widespread ParB family, Noc and Spo0J are required to prevent Z ring assembly over the bacterial nucleoid and help fine tune the assembly of the Z ring at midcell during the cell cycle.
© 2019 The Authors. Molecular Microbiology Published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

References
-
- Autret, S. , Nair, R. and Errington, J. (2001) Genetic analysis of the chromosome segregation protein Spo0J of Bacillus subtilis: evidence for separate domains involved in DNA binding and interactions with Soj protein. Molecular Microbiology, 41, 743–755. - PubMed
-
- Bazill, G.W. and Gross, J.D. (1972) Effect of 6‐(p‐hydroxyphenyl)‐azouracil on B. subtilis DNA polymerases. Nat New Biol, 240, 82–83. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
