

Structure and Mechanism of Acetylation by the N-Terminal Dual Enzyme NatA/Naa50 Complex
- PMID: 31155310
- PMCID: PMC6610660
- DOI: 10.1016/j.str.2019.04.014
Structure and Mechanism of Acetylation by the N-Terminal Dual Enzyme NatA/Naa50 Complex
Abstract
NatA co-translationally acetylates the N termini of over 40% of eukaryotic proteins and can associate with another catalytic subunit, Naa50, to form a ternary NatA/Naa50 dual enzyme complex (also called NatE). The molecular basis of association between Naa50 and NatA and the mechanism for how their association affects their catalytic activities in yeast and human are poorly understood. Here, we determined the X-ray crystal structure of yeast NatA/Naa50 as a scaffold to understand coregulation of NatA/Naa50 activity in both yeast and human. We find that Naa50 makes evolutionarily conserved contacts to both the Naa10 and Naa15 subunits of NatA. These interactions promote catalytic crosstalk within the human complex, but do so to a lesser extent in the yeast complex, where Naa50 activity is compromised. These studies have implications for understanding the role of the NatA/Naa50 complex in modulating the majority of the N-terminal acetylome in diverse species.
Keywords: N-terminal acetylation; Naa50; NatA; NatE; X-ray crystallography; protein complex.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interest.
Figures
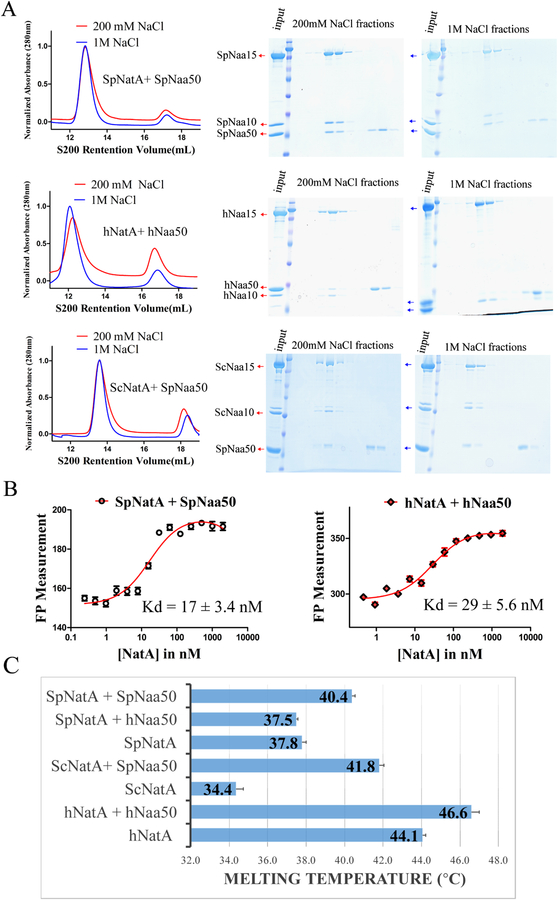
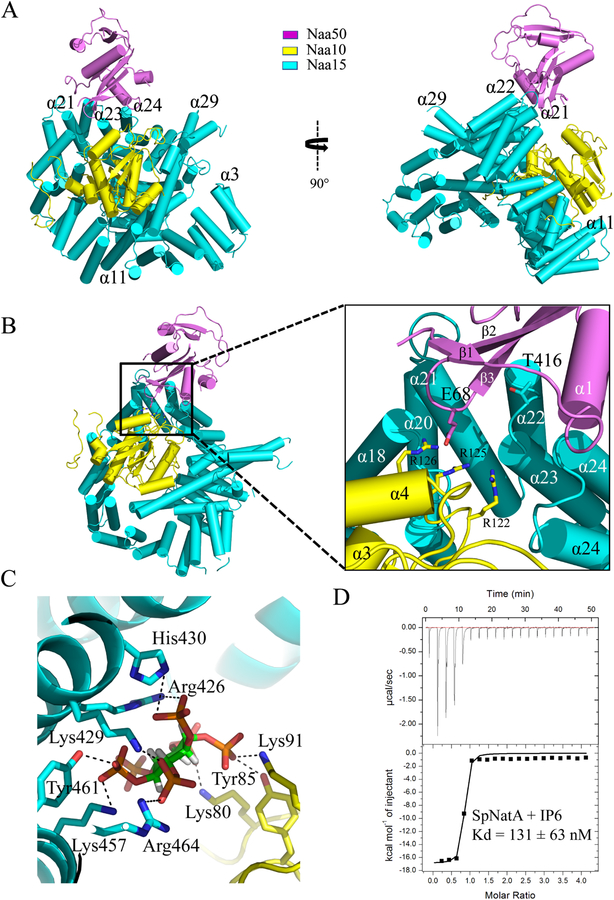
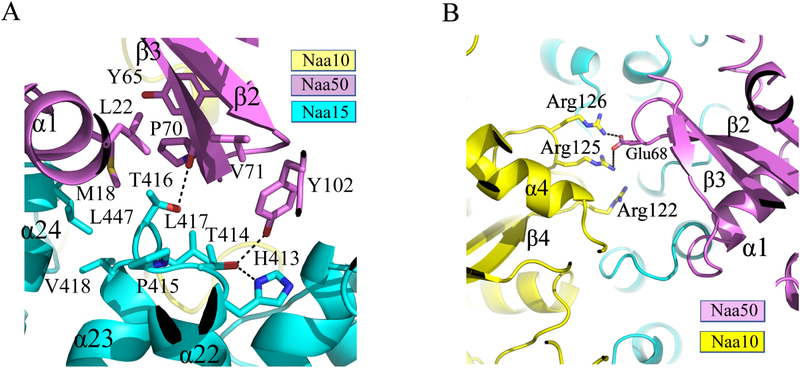
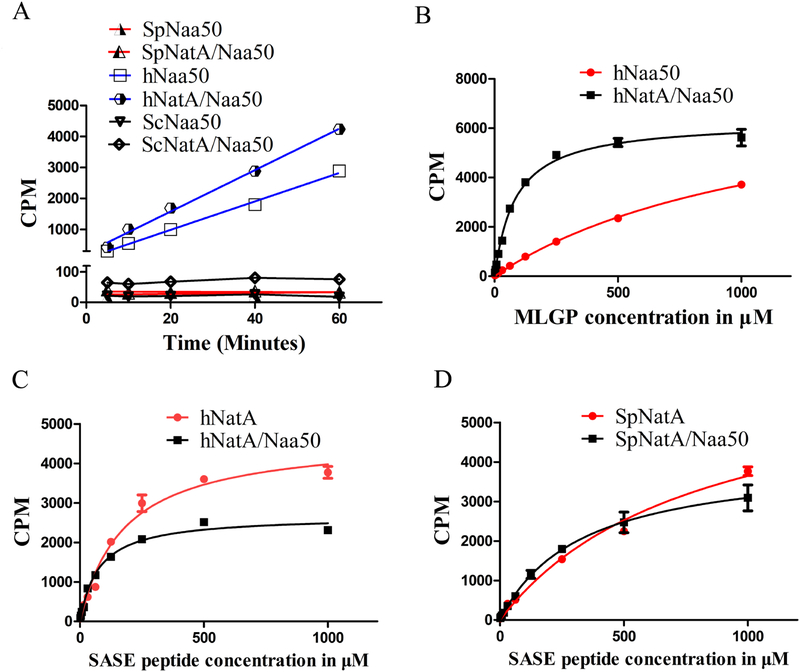
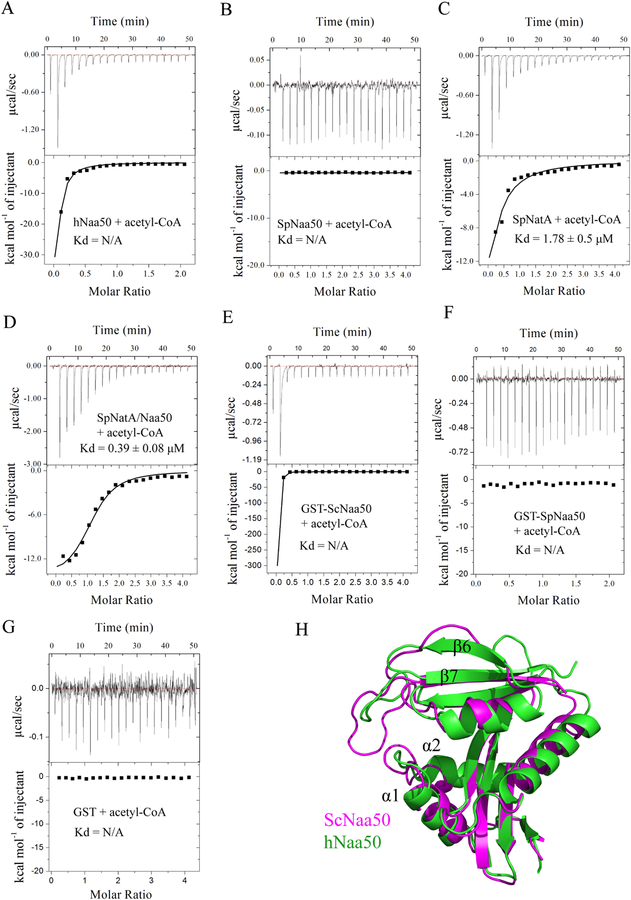
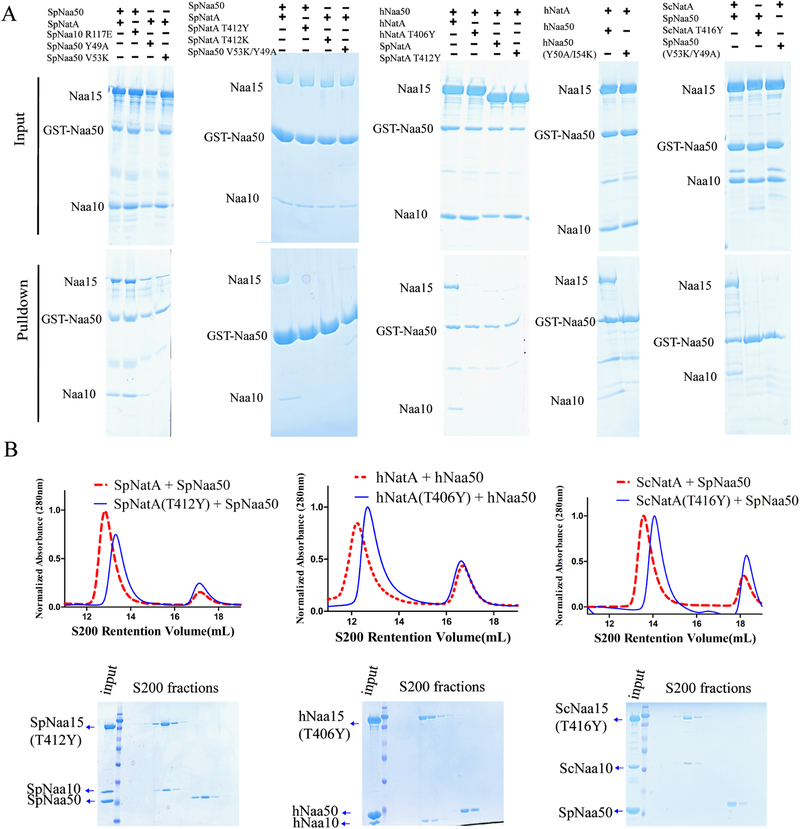
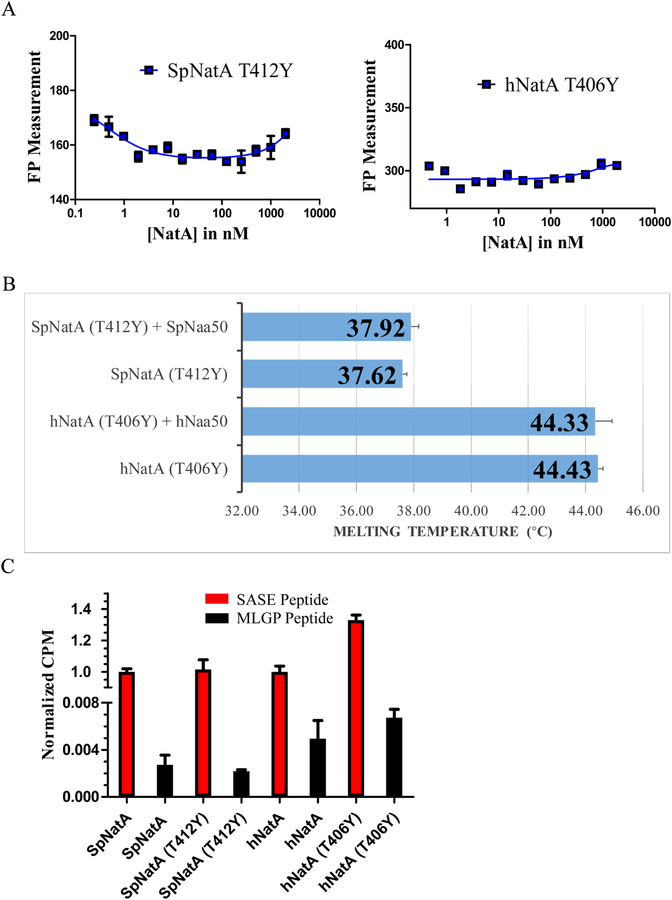
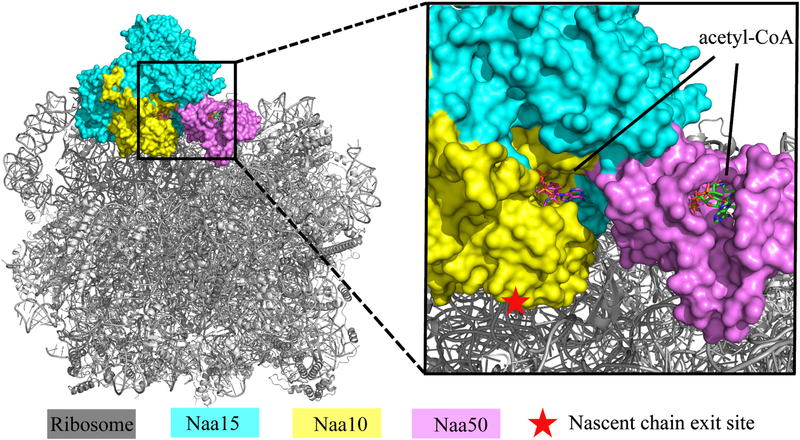
Comment in
-
From Molecular Understanding to Organismal Biology of N-Terminal Acetyltransferases.Structure. 2019 Jul 2;27(7):1053-1055. doi: 10.1016/j.str.2019.06.002. Structure. 2019. PMID: 31269458 Free PMC article.
References
-
- ABUL KALAM S & SANCHITA B 2012. IP6 (Inositol Hexaphosphate) as a Signaling Molecule. Current Signal Transduction Therapy, 7, 289–304.
-
- ADAMS PD, AFONINE PV, BUNKOCZI G, CHEN VB, DAVIS IW, ECHOLS N, HEADD JJ, HUNG L-W, KAPRAL GJ,GROSSE-KUNSTLEVE RW, MCCOY AJ, MORIARTY NW, OEFFNER R, READ RJ, RICHARDSON DC, RICHARDSON JS, TERWILLIGER TC & ZWART PH 2010. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallographica Section D, 66, 213–221. - PMC - PubMed
-
- AKSNES H, DRAZIC A, MARIE M & ARNESEN T 2016. First Things First: Vital Protein Marks by N-Terminal Acetyltransferases. Trends in Biochemical Sciences, 41, 746–760. - PubMed
-
- AKSNES H, VAN DAMME P, GORIS M, STARHEIM KK, MARIE M, STOVE SI, HOEL C, KALVIK TV, HOLE K, GLOMNES N, FURNES C, LJOSTVEIT S, ZIEGLER M, NIERE M, GEVAERT K & ARNESEN T 2015. An organellar nalpha-acetyltransferase, naa60, acetylates cytosolic N termini of transmembrane proteins and maintains Golgi integrity. Cell Rep, 10, 1362–74. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
