

Reconstruction and Simulation of a Scaffold Model of the Cerebellar Network
- PMID: 31156416
- PMCID: PMC6530631
- DOI: 10.3389/fninf.2019.00037
Reconstruction and Simulation of a Scaffold Model of the Cerebellar Network
Erratum in
-
Corrigendum: Reconstruction and Simulation of a Scaffold Model of the Cerebellar Network.Front Neuroinform. 2019 Jul 10;13:51. doi: 10.3389/fninf.2019.00051. eCollection 2019. Front Neuroinform. 2019. PMID: 31354466 Free PMC article.
Abstract
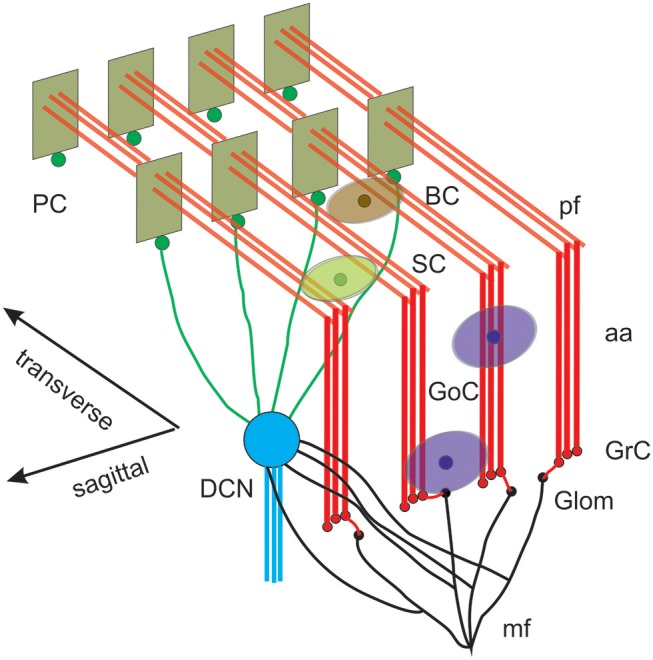
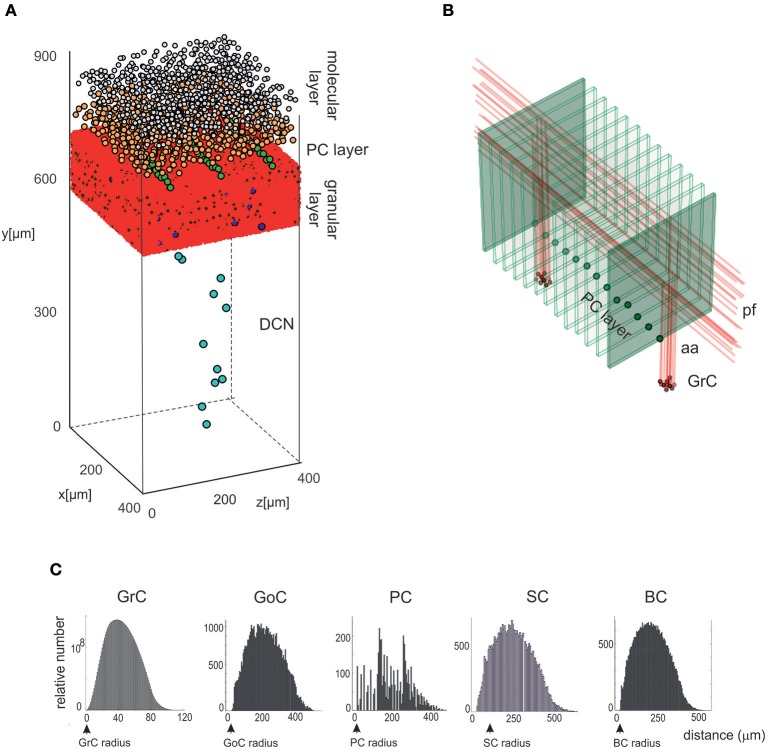
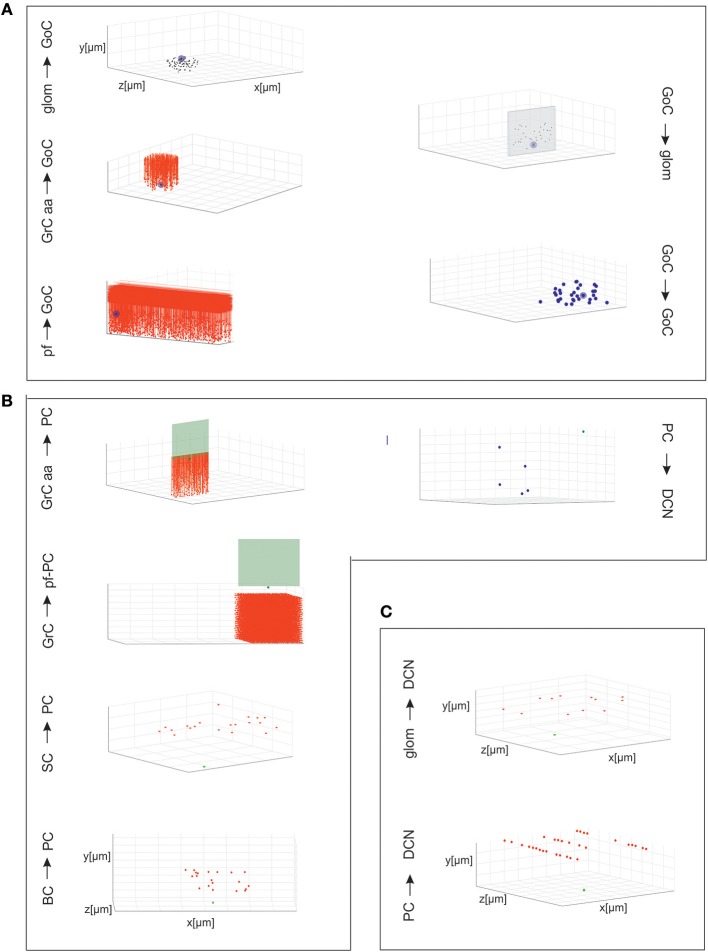
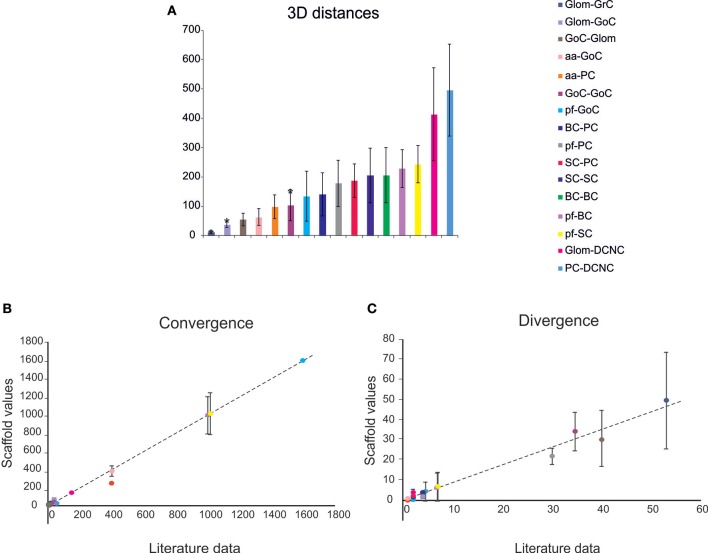
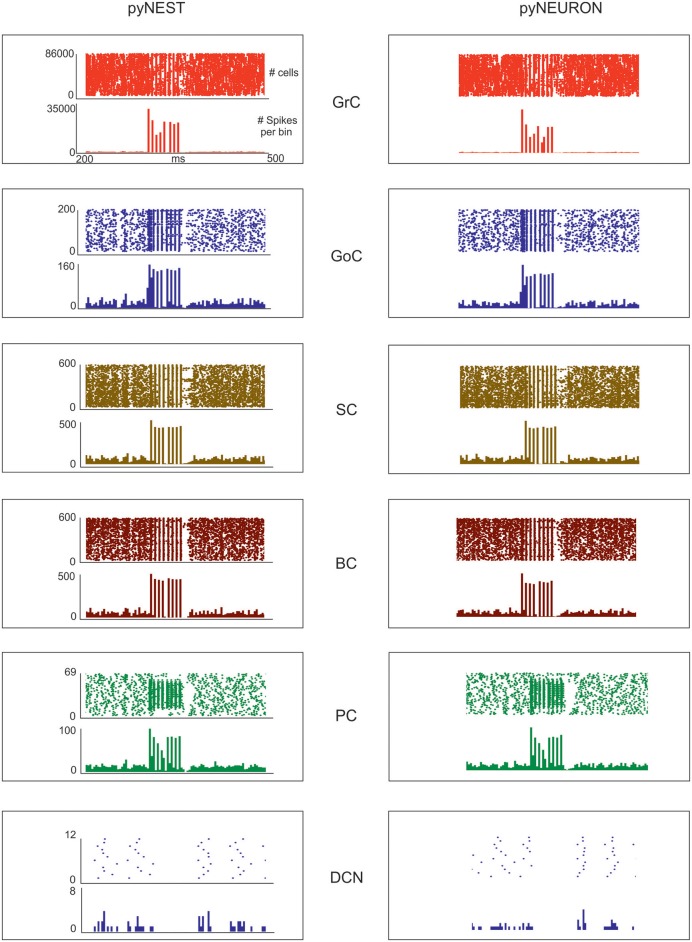
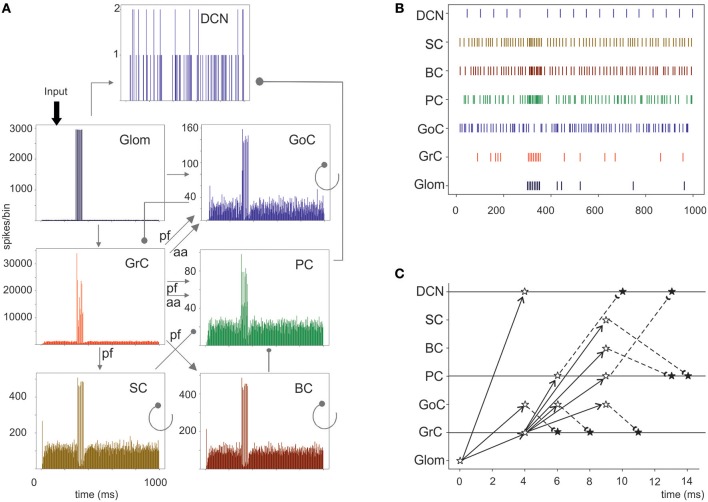
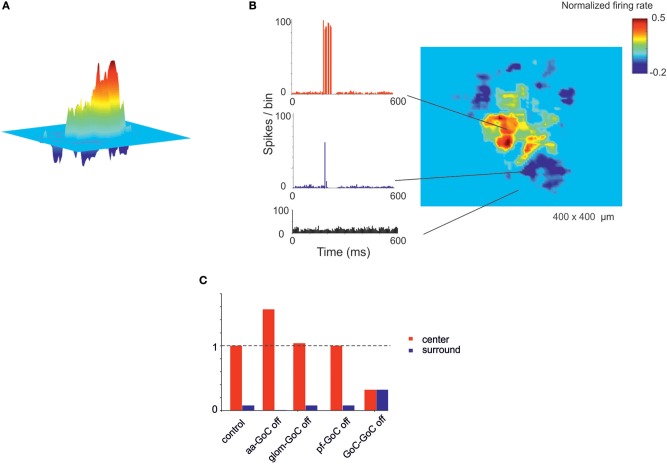
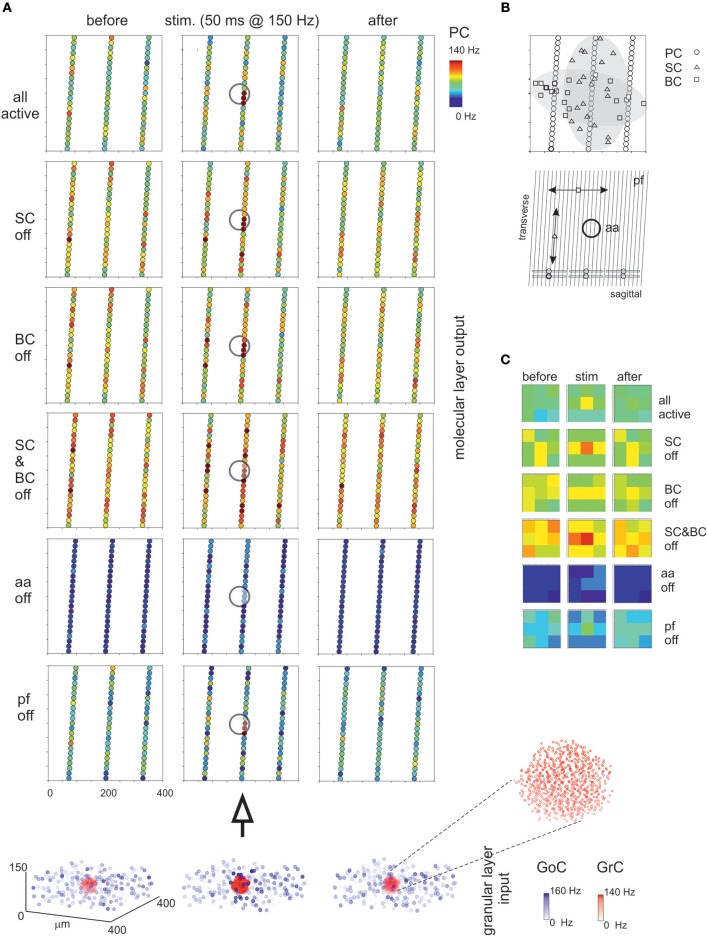
Reconstructing neuronal microcircuits through computational models is fundamental to simulate local neuronal dynamics. Here a scaffold model of the cerebellum has been developed in order to flexibly place neurons in space, connect them synaptically, and endow neurons and synapses with biologically-grounded mechanisms. The scaffold model can keep neuronal morphology separated from network connectivity, which can in turn be obtained from convergence/divergence ratios and axonal/dendritic field 3D geometries. We first tested the scaffold on the cerebellar microcircuit, which presents a challenging 3D organization, at the same time providing appropriate datasets to validate emerging network behaviors. The scaffold was designed to integrate the cerebellar cortex with deep cerebellar nuclei (DCN), including different neuronal types: Golgi cells, granule cells, Purkinje cells, stellate cells, basket cells, and DCN principal cells. Mossy fiber inputs were conveyed through the glomeruli. An anisotropic volume (0.077 mm3) of mouse cerebellum was reconstructed, in which point-neuron models were tuned toward the specific discharge properties of neurons and were connected by exponentially decaying excitatory and inhibitory synapses. Simulations using both pyNEST and pyNEURON showed the emergence of organized spatio-temporal patterns of neuronal activity similar to those revealed experimentally in response to background noise and burst stimulation of mossy fiber bundles. Different configurations of granular and molecular layer connectivity consistently modified neuronal activation patterns, revealing the importance of structural constraints for cerebellar network functioning. The scaffold provided thus an effective workflow accounting for the complex architecture of the cerebellar network. In principle, the scaffold can incorporate cellular mechanisms at multiple levels of detail and be tuned to test different structural and functional hypotheses. A future implementation using detailed 3D multi-compartment neuron models and dynamic synapses will be needed to investigate the impact of single neuron properties on network computation.
Keywords: Python; cerebellum; computational spiking models; connectome; pyNEST; pyNEURON.
Figures
References
LinkOut - more resources
Full Text Sources
Miscellaneous