

Emergence of three-dimensional order and structure in growing biofilms
- PMID: 31156716
- PMCID: PMC6544526
- DOI: 10.1038/s41567-018-0356-9
Emergence of three-dimensional order and structure in growing biofilms
Abstract
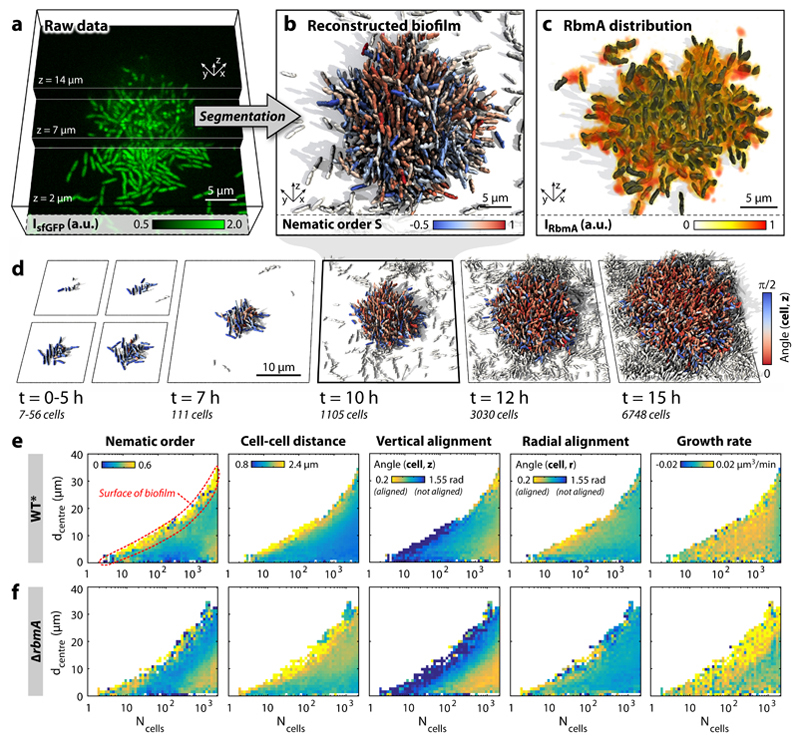
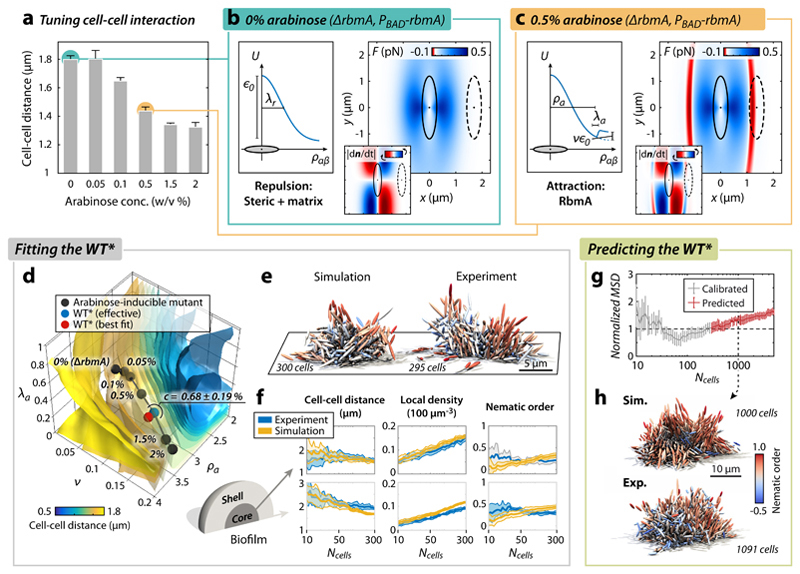
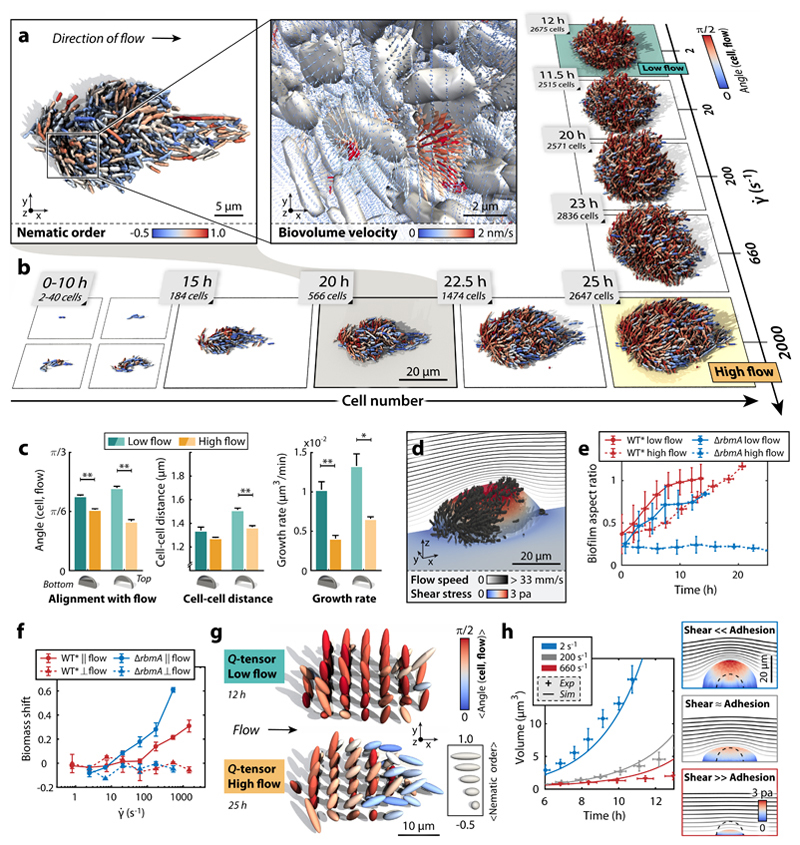
Surface-attached bacterial biofilms are self-replicating active liquid crystals and the dominant form of bacterial life on earth (1-4). In conventional liquid crystals and solid-state materials, the interaction potentials between the molecules that comprise the system determine the material properties. However, for growth-active biofilms it is unclear whether potential-based descriptions can account for the experimentally observed morphologies, and which potentials would be relevant. Here, we overcame previous limitations of single-cell imaging techniques (5,6) to reconstruct and track all individual cells inside growing three-dimensional (3D) biofilms with up to 10,000 individuals. Based on these data, we identify, constrain, and provide a microscopic basis for an effective cell-cell interaction potential, which captures and predicts the growth dynamics, emergent architecture, and local liquid crystalline order of Vibrio cholerae biofilms. Furthermore, we show how external fluid flows control the microscopic structure and 3D morphology of biofilms. Our analysis implies that local cellular order and global biofilm architecture in these active bacterial communities can arise from mechanical cell-cell interactions, which cells can modulate by regulating the production of particular matrix components. These results establish an experimentally validated foundation for improved continuum theories of active matter and thereby contribute to solving the important problem of controlling biofilm growth.
Conflict of interest statement
Competing financial interests The authors declare no competing financial interest.
Figures
References
-
- Hagan MF, Baskaran A. Emergent self-organization in active materials. Curr Opin Cell Biol. 2016;38:74–80. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources