

Priming of Hypothalamic Ghrelin Signaling and Microglia Activation Exacerbate Feeding in Rats' Offspring Following Maternal Overnutrition
- PMID: 31159189
- PMCID: PMC6627862
- DOI: 10.3390/nu11061241
Priming of Hypothalamic Ghrelin Signaling and Microglia Activation Exacerbate Feeding in Rats' Offspring Following Maternal Overnutrition
Abstract
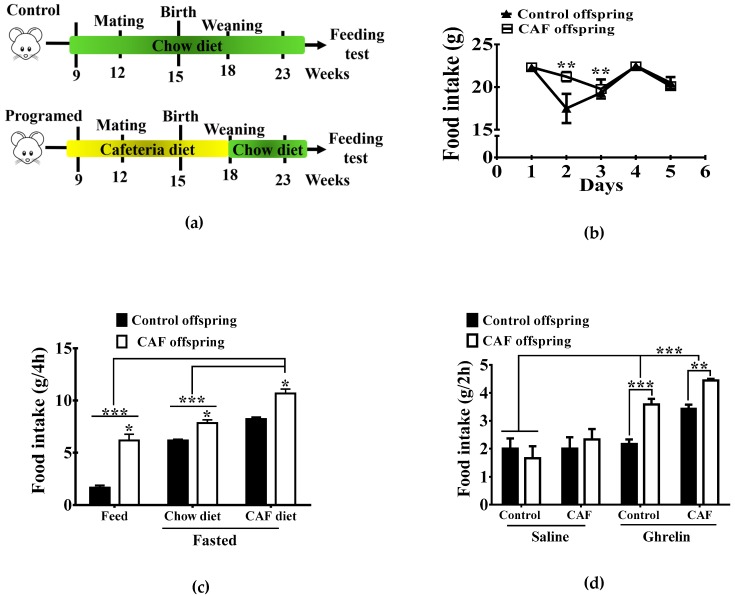
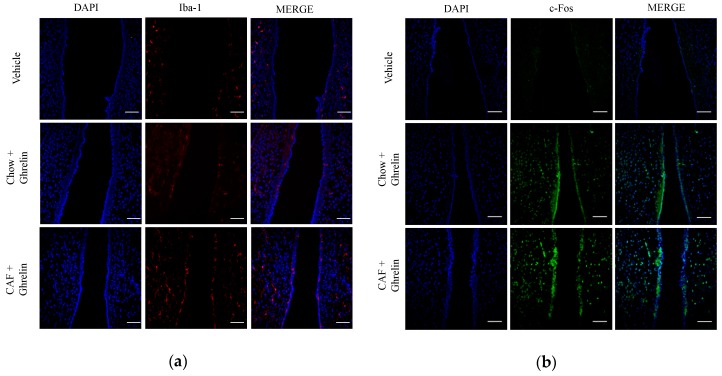
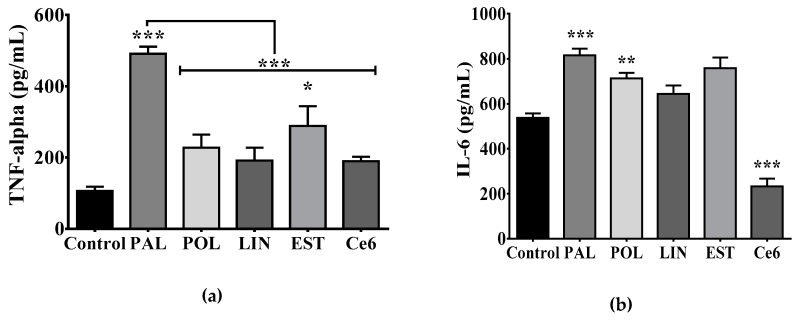
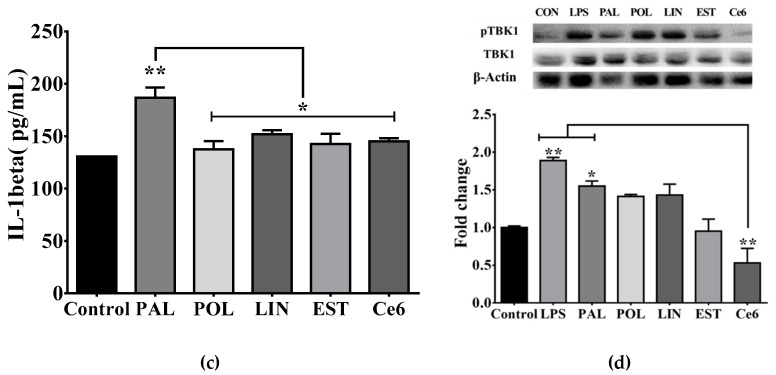
Maternal overnutrition during pregnancy leads to metabolic alterations, including obesity, hyperphagia, and inflammation in the offspring. Nutritional priming of central inflammation and its role in ghrelin sensitivity during fed and fasted states have not been analyzed. The current study aims to identify the effect of maternal programming on microglia activation and ghrelin-induced activation of hypothalamic neurons leading to food intake response. We employed a nutritional programming model exposing female Wistar rats to a cafeteria diet (CAF) from pre-pregnancy to weaning. Food intake in male offspring was determined daily after fasting and subcutaneous injection of ghrelin. Hypothalamic ghrelin sensitivity and microglia activation was evaluated using immunodetection for Iba-1 and c-Fos markers, and Western blot for TBK1 signaling. Release of TNF-alpha, IL-6, and IL-1β after stimulation with palmitic, oleic, linoleic acid, or C6 ceramide in primary microglia culture were quantified using ELISA. We found that programmed offspring by CAF diet exhibits overfeeding after fasting and peripheral ghrelin administration, which correlates with an increase in the hypothalamic Iba-1 microglia marker and c-Fos cell activation. Additionally, in contrast to oleic, linoleic, or C6 ceramide stimulation in primary microglia culture, stimulation with palmitic acid for 24 h promotes TNF-alpha, IL-6, and IL-1β release and TBK1 activation. Notably, intracerebroventricular (i.c.v.) palmitic acid or LPS inoculation for five days promotes daily increase in food intake and food consumption after ghrelin administration. Finally, we found that i.c.v. palmitic acid substantially activates hypothalamic Iba-1 microglia marker and c-Fos. Together, our results suggest that maternal nutritional programing primes ghrelin sensitivity and microglia activation, which potentially might mirror hypothalamic administration of the saturated palmitic acid.
Keywords: ghrelin; hypothalamic inflammation; microglia; nutritional programing.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
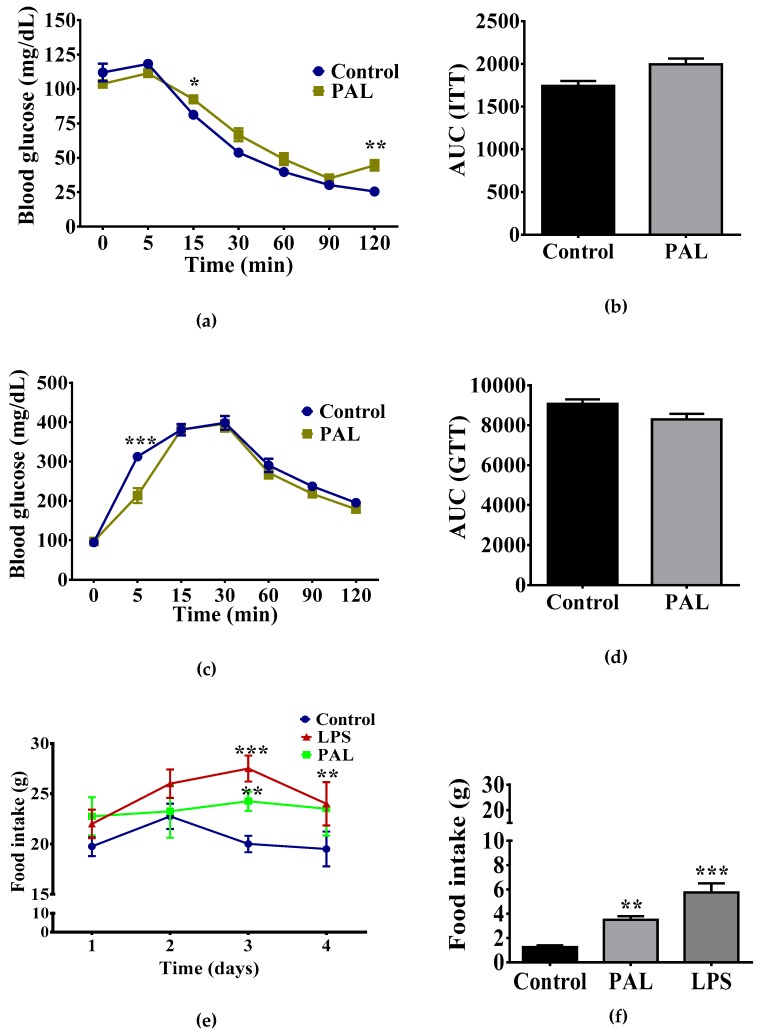
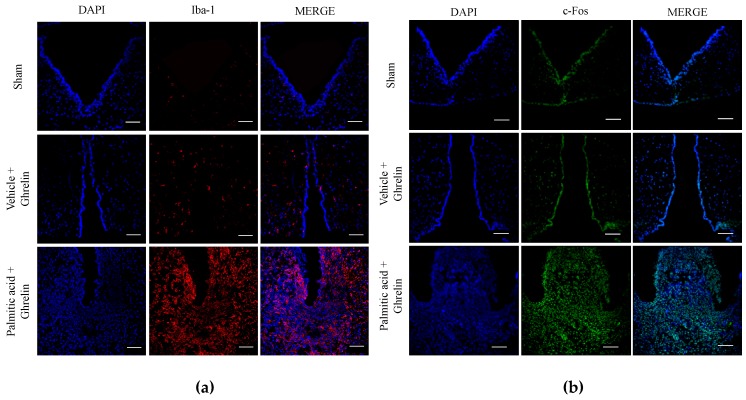
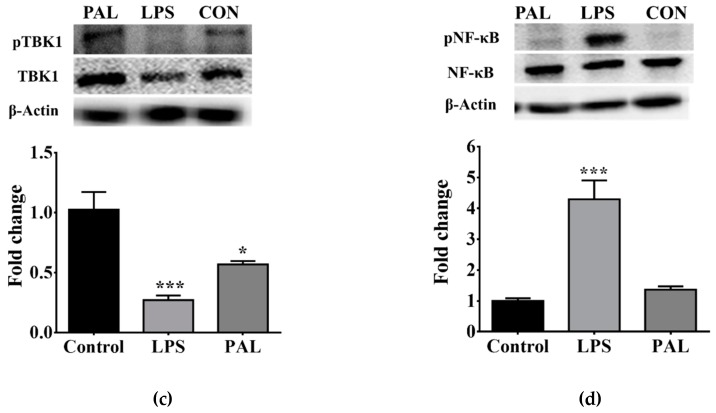
References
-
- Alfaradhi M.Z., Kusinski L.C., Fernandez-Twinn D.S., Pantaleão L.C., Carr S.K., Ferland-McCollough D., Yeo G.S.H., Bushell M., Ozanne E.S. Maternal Obesity in Pregnancy Developmentally Programs Adipose Tissue Inflammation in Young, Lean Male Mice Offspring. Endocrinology. 2016;157:4246–4256. doi: 10.1210/en.2016-1314. - DOI - PMC - PubMed
-
- Frihauf J.B., Fekete É.M., Nagy T.R., Levin B.E., Zorrilla E.P. Maternal Western diet increases adiposity even in male offspring of obesity-resistant rat dams: Early endocrine risk markers. Am. J. Physiol. Integr. Comp. Physiol. 2016;311:R1045–R1059. doi: 10.1152/ajpregu.00023.2016. - DOI - PMC - PubMed
-
- Cardenas-Perez R.E., Fuentes-Mera L., de la Garza A.L., Torre-Villalvazo I., Reyes-Castro L.A., Rodriguez-Rocha H., Garcia-Garcia A., Corona-Castillo J.C., Tovar A.R., Zambrano E., et al. Maternal overnutrition by hypercaloric diets programs hypothalamic mitochondrial fusion and metabolic dysfunction in rat male offspring. Nutr. Metab. (London) 2018;15:38. doi: 10.1186/s12986-018-0279-6. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
