

Role of tumor and host autophagy in cancer metabolism
- PMID: 31160394
- PMCID: PMC6546058
- DOI: 10.1101/gad.325514.119
Role of tumor and host autophagy in cancer metabolism
Abstract
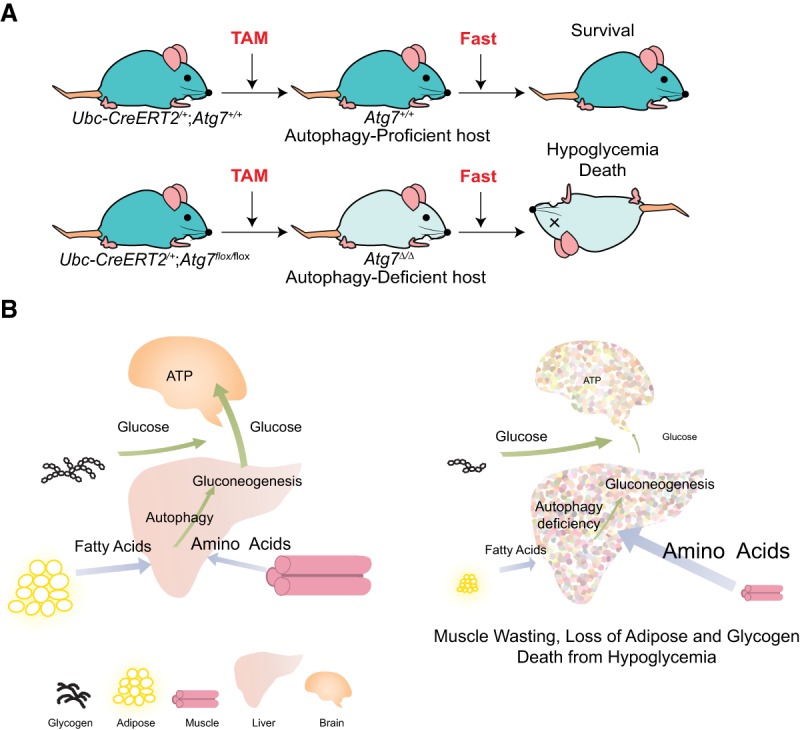
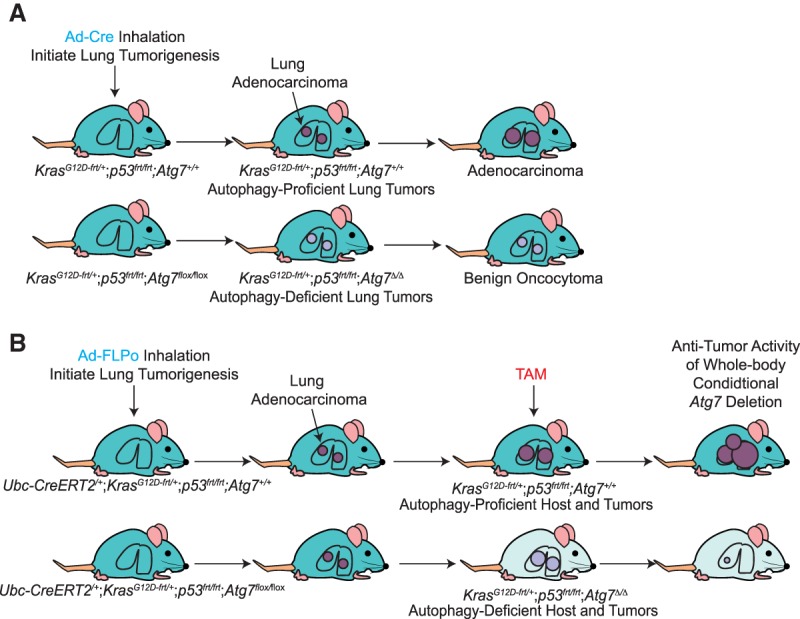
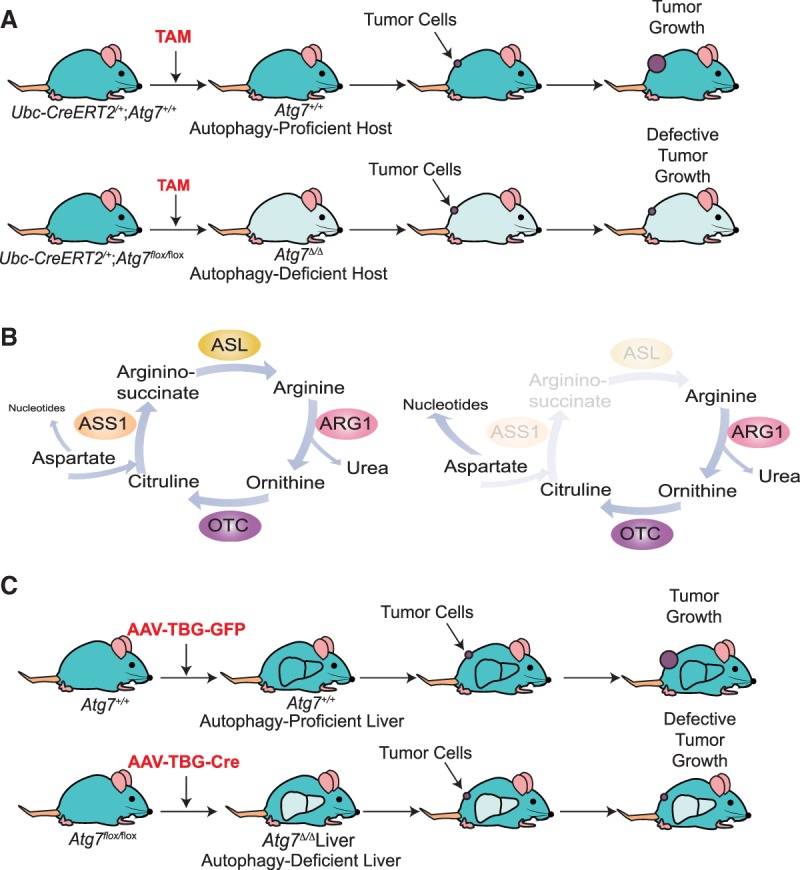
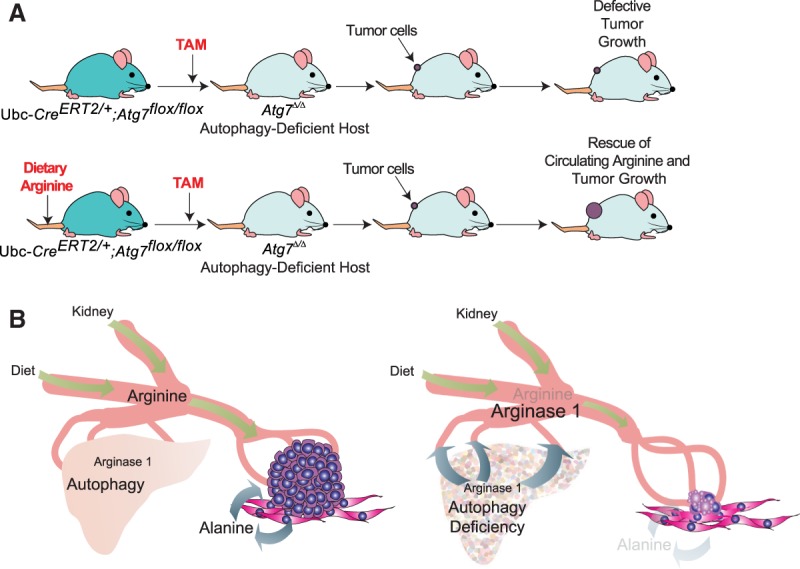
Macroautophagy (referred to here as autophagy) degrades and recycles cytoplasmic constituents to sustain cellular and mammalian metabolism and survival during starvation. Deregulation of autophagy is involved in numerous diseases, such as cancer. Cancers up-regulate autophagy and depend on it for survival, growth, and malignancy in a tumor cell-autonomous fashion. Recently, it has become apparent that autophagy in host tissues as well as the tumor cells themselves contribute to tumor growth. Understanding how autophagy regulates metabolism and tumor growth has revealed new essential tumor nutrients, where they come from, and how they are supplied and used, which can now be targeted for cancer therapy.
Keywords: autophagy; cancer; metabolism.
© 2019 Poillet-Perez and White; Published by Cold Spring Harbor Laboratory Press.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources