

Structural basis for substrate gripping and translocation by the ClpB AAA+ disaggregase
- PMID: 31160557
- PMCID: PMC6546751
- DOI: 10.1038/s41467-019-10150-y
Structural basis for substrate gripping and translocation by the ClpB AAA+ disaggregase
Abstract
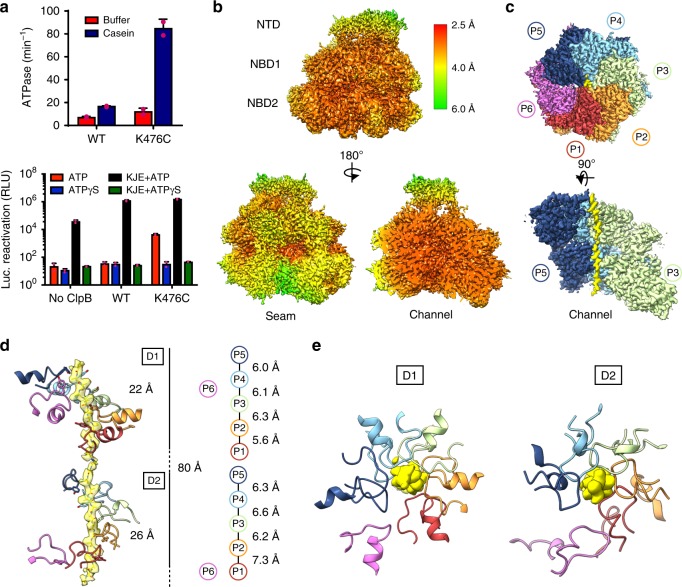
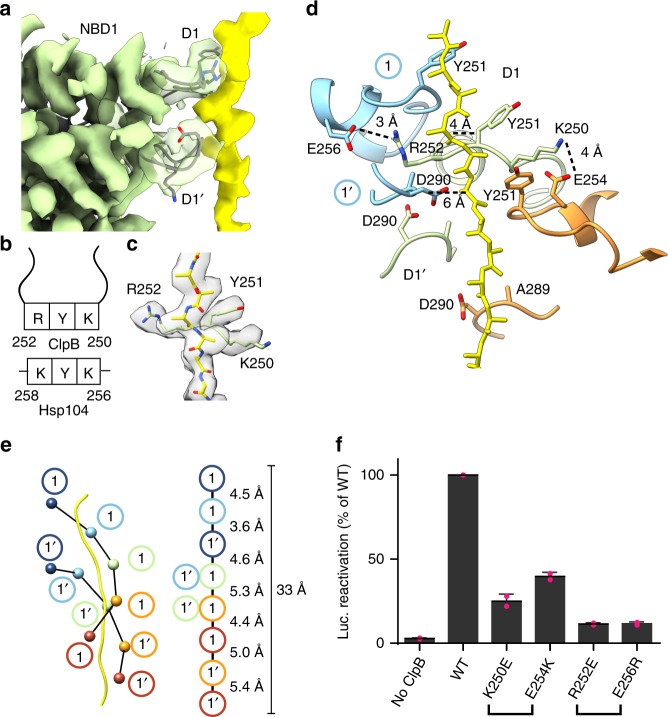
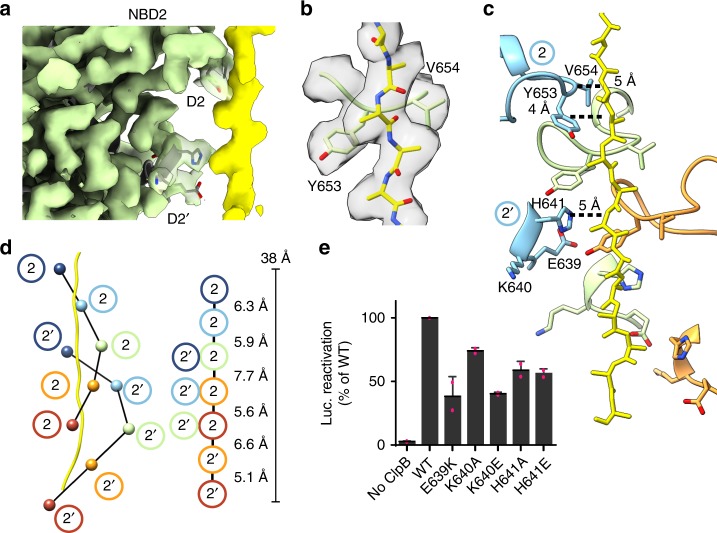
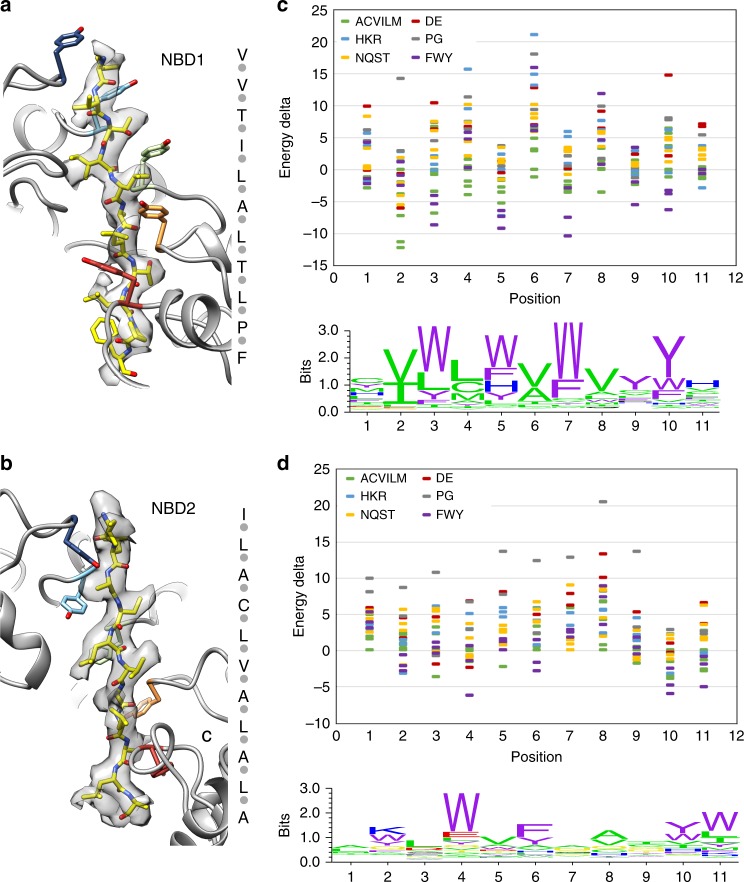
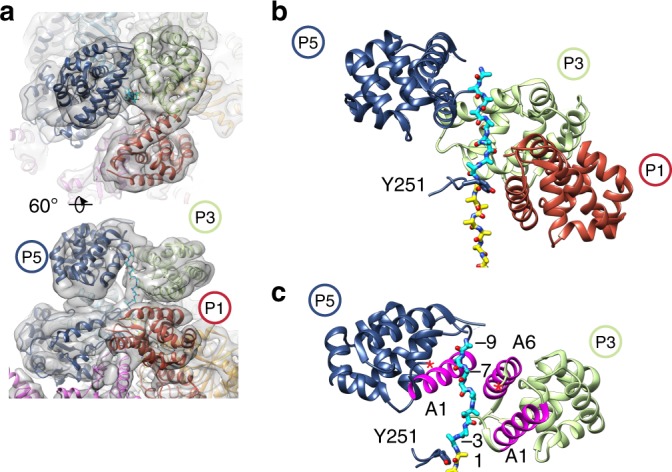
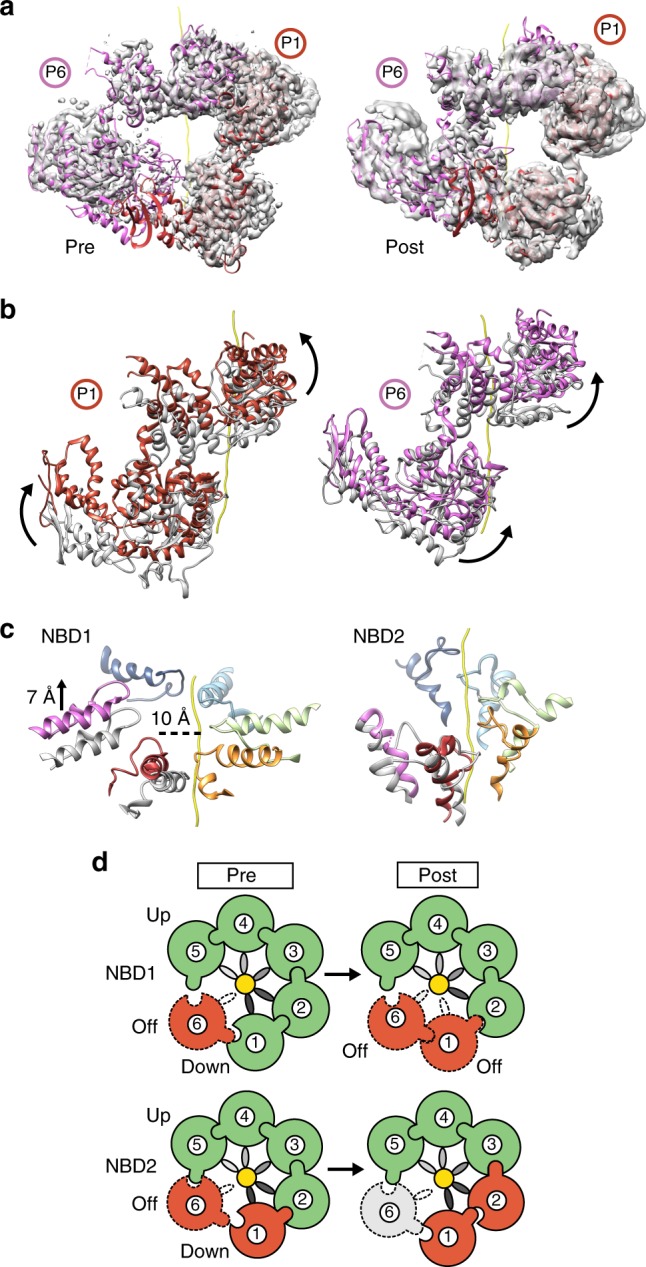
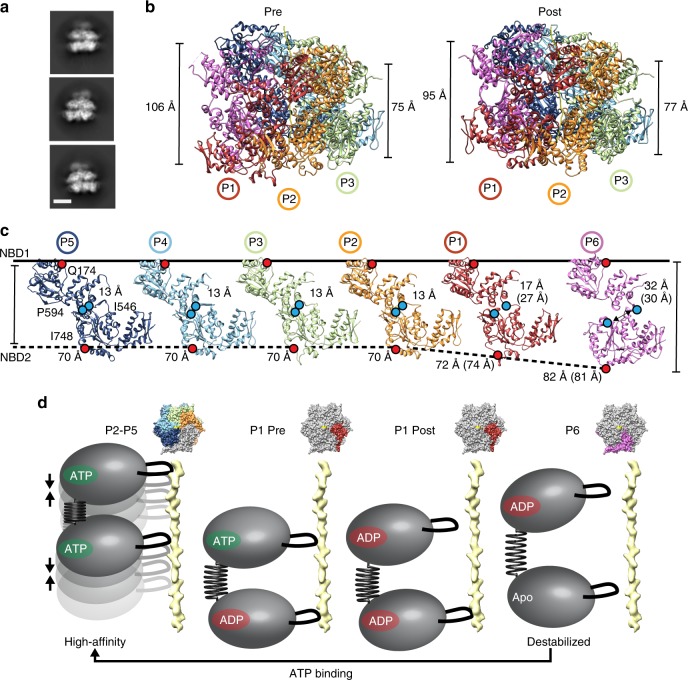
Bacterial ClpB and yeast Hsp104 are homologous Hsp100 protein disaggregases that serve critical functions in proteostasis by solubilizing protein aggregates. Two AAA+ nucleotide binding domains (NBDs) power polypeptide translocation through a central channel comprised of a hexameric spiral of protomers that contact substrate via conserved pore-loop interactions. Here we report cryo-EM structures of a hyperactive ClpB variant bound to the model substrate, casein in the presence of slowly hydrolysable ATPγS, which reveal the translocation mechanism. Distinct substrate-gripping interactions are identified for NBD1 and NBD2 pore loops. A trimer of N-terminal domains define a channel entrance that binds the polypeptide substrate adjacent to the topmost NBD1 contact. NBD conformations at the seam interface reveal how ATP hydrolysis-driven substrate disengagement and re-binding are precisely tuned to drive a directional, stepwise translocation cycle.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Two-Step Activation Mechanism of the ClpB Disaggregase for Sequential Substrate Threading by the Main ATPase Motor.Cell Rep. 2019 Jun 18;27(12):3433-3446.e4. doi: 10.1016/j.celrep.2019.05.075. Cell Rep. 2019. PMID: 31216466 Free PMC article.
-
ATP hydrolysis-coupled peptide translocation mechanism of Mycobacterium tuberculosis ClpB.Proc Natl Acad Sci U S A. 2018 Oct 9;115(41):E9560-E9569. doi: 10.1073/pnas.1810648115. Epub 2018 Sep 26. Proc Natl Acad Sci U S A. 2018. PMID: 30257943 Free PMC article.
-
Ratchet-like polypeptide translocation mechanism of the AAA+ disaggregase Hsp104.Science. 2017 Jul 21;357(6348):273-279. doi: 10.1126/science.aan1052. Epub 2017 Jun 15. Science. 2017. PMID: 28619716 Free PMC article.
-
Spiraling in Control: Structures and Mechanisms of the Hsp104 Disaggregase.Cold Spring Harb Perspect Biol. 2019 Aug 1;11(8):a034033. doi: 10.1101/cshperspect.a034033. Cold Spring Harb Perspect Biol. 2019. PMID: 30745294 Free PMC article. Review.
-
Molecular chaperones: structure of a protein disaggregase.Curr Biol. 2004 Jan 20;14(2):R78-80. doi: 10.1016/j.cub.2003.12.051. Curr Biol. 2004. PMID: 14738756 Review.
Cited by
-
Unique structural features govern the activity of a human mitochondrial AAA+ disaggregase, Skd3.Cell Rep. 2022 Sep 27;40(13):111408. doi: 10.1016/j.celrep.2022.111408. Cell Rep. 2022. PMID: 36170828 Free PMC article.
-
Protein quality control: from mechanism to disease : EMBO Workshop, Costa de la Calma (Mallorca), Spain, April 28 - May 03, 2019.Cell Stress Chaperones. 2019 Nov;24(6):1013-1026. doi: 10.1007/s12192-019-01040-9. Epub 2019 Nov 11. Cell Stress Chaperones. 2019. PMID: 31713048 Free PMC article.
-
Recent structural insights into the mechanism of ClpP protease regulation by AAA+ chaperones and small molecules.J Biol Chem. 2022 May;298(5):101781. doi: 10.1016/j.jbc.2022.101781. Epub 2022 Mar 2. J Biol Chem. 2022. PMID: 35245501 Free PMC article. Review.
-
Conformational plasticity of the ClpAP AAA+ protease couples protein unfolding and proteolysis.Nat Struct Mol Biol. 2020 May;27(5):406-416. doi: 10.1038/s41594-020-0409-5. Epub 2020 Apr 20. Nat Struct Mol Biol. 2020. PMID: 32313240 Free PMC article.
-
The unfoldase ClpC1 of Mycobacterium tuberculosis regulates the expression of a distinct subset of proteins having intrinsically disordered termini.J Biol Chem. 2020 Jul 10;295(28):9455-9473. doi: 10.1074/jbc.RA120.013456. Epub 2020 May 14. J Biol Chem. 2020. PMID: 32409584 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
