

Defective viral genomes are key drivers of the virus-host interaction
- PMID: 31160826
- PMCID: PMC7097797
- DOI: 10.1038/s41564-019-0465-y
Defective viral genomes are key drivers of the virus-host interaction
Abstract
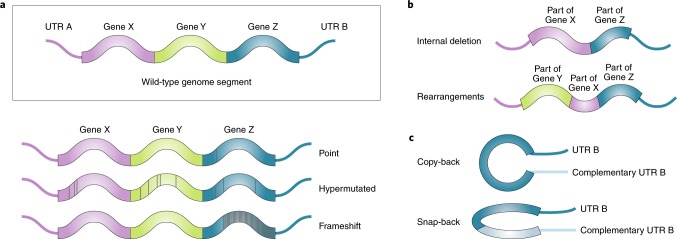
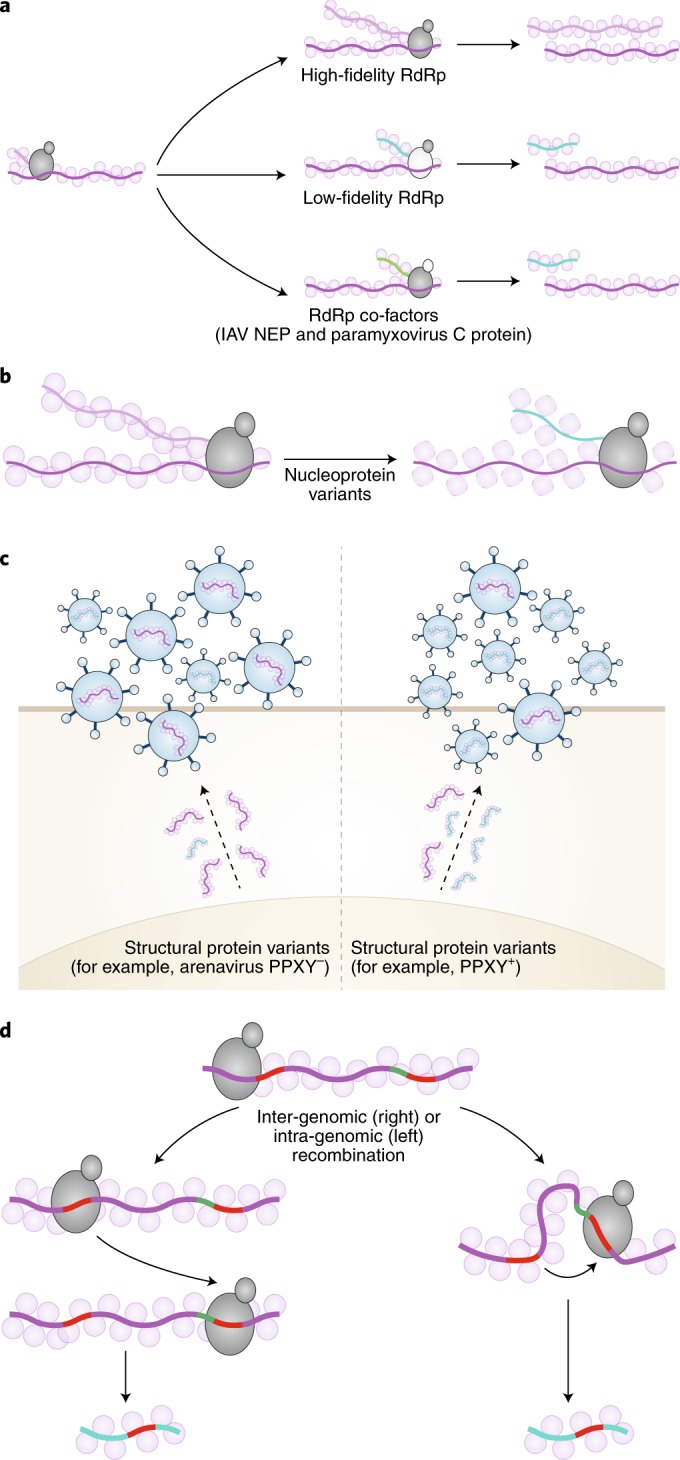
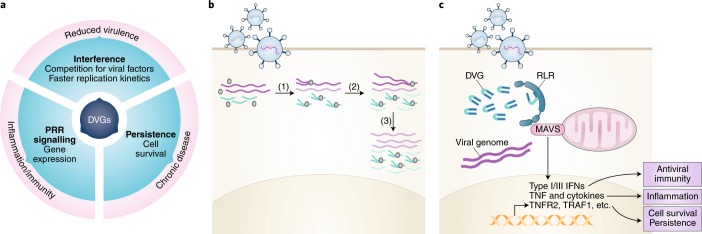
Viruses survive often harsh host environments, yet we know little about the strategies they utilize to adapt and subsist given their limited genomic resources. We are beginning to appreciate the surprising versatility of viral genomes and how replication-competent and -defective virus variants can provide means for adaptation, immune escape and virus perpetuation. This Review summarizes current knowledge of the types of defective viral genomes generated during the replication of RNA viruses and the functions that they carry out. We highlight the universality and diversity of defective viral genomes during infections and discuss their predicted role in maintaining a fit virus population, their impact on human and animal health, and their potential to be harnessed as antiviral tools.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Poirier EZ, Vignuzzi M. Virus population dynamics during infection. Curr. Opin. Virol. 2017;23:82–87. - PubMed
-
- Von Magnus P, Gard S. Studies on interference in experimental influenza. Ark. Kemi. Mineral. Geol. 1947;24:4.
-
- Huang AS, Baltimore D. Defective viral particles and viral disease processes. Nature. 1970;226:325–327. - PubMed
-
- Barrett AD, Dimmock NJ. Modulation of Semliki Forest virus-induced infection of mice by defective-interfering virus. J. Infect. Dis. 1984;150:98–104. - PubMed
-
- Rabinowitz SG, Huprikar J. The influence of defective-interfering particles of the PR-8 strain of influenza A virus on the pathogenesis of pulmonary infection in mice. J. Infect. Dis. 1979;140:305–315. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
