

Stress-Induced Changes in the Lipid Microenvironment of β-(1,3)-d-Glucan Synthase Cause Clinically Important Echinocandin Resistance in Aspergillus fumigatus
- PMID: 31164462
- PMCID: PMC6550521
- DOI: 10.1128/mBio.00779-19
Stress-Induced Changes in the Lipid Microenvironment of β-(1,3)-d-Glucan Synthase Cause Clinically Important Echinocandin Resistance in Aspergillus fumigatus
Abstract
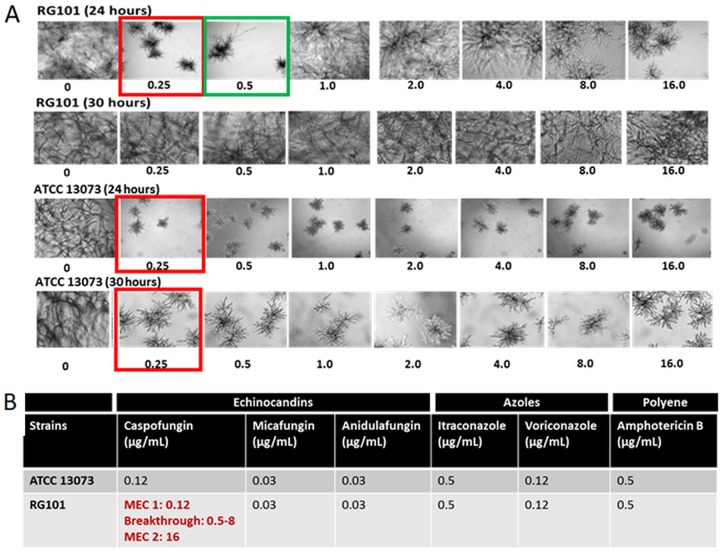
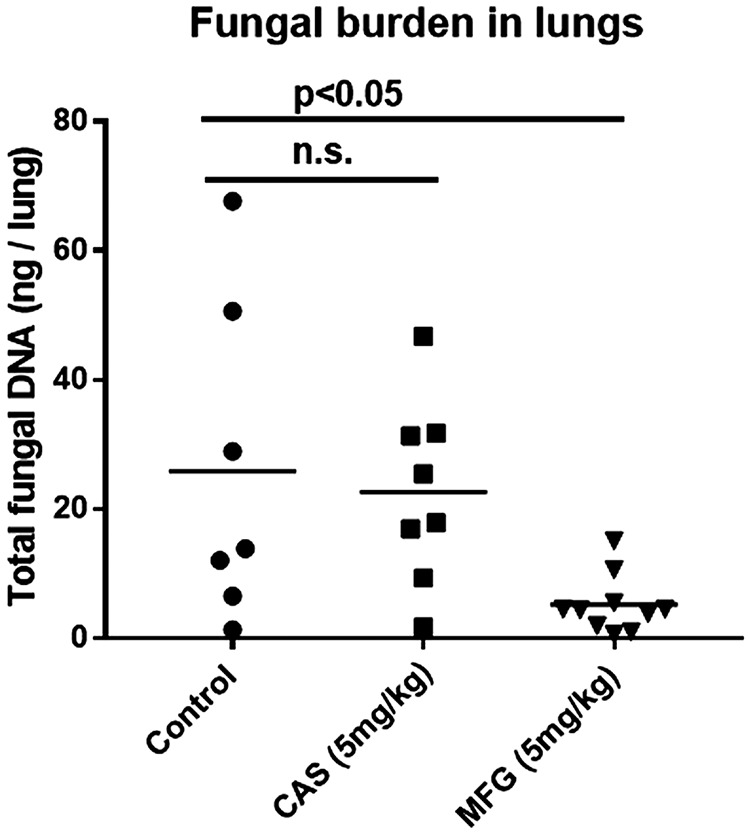
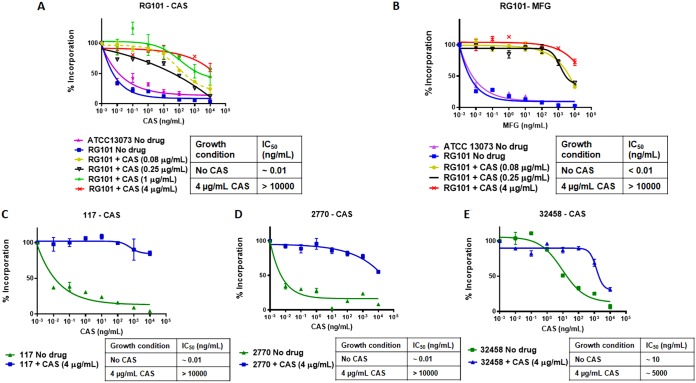
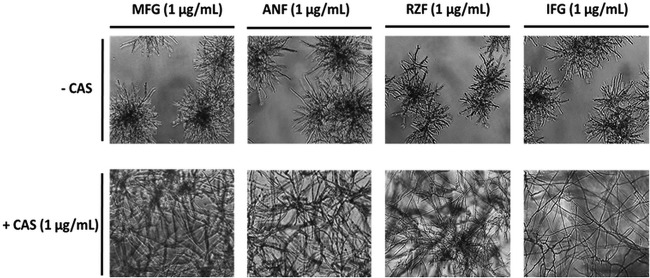
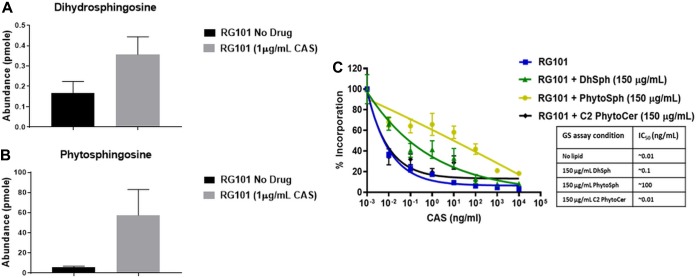
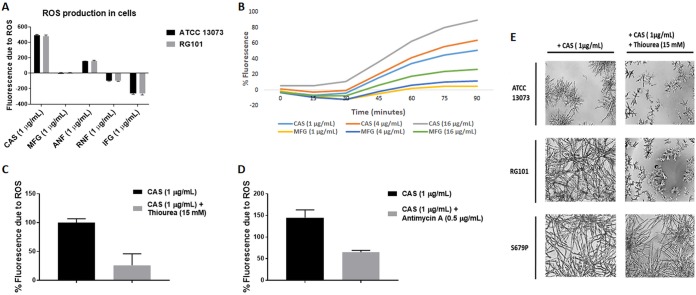
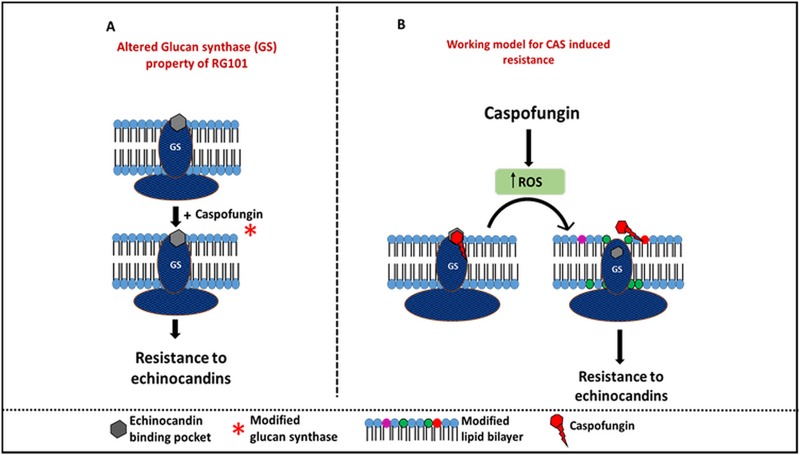
Aspergillus fumigatus is a leading cause of invasive fungal infections. Resistance to first-line triazole antifungals has led to therapy with echinocandin drugs. Recently, we identified several high-minimum-effective-concentration (MEC) A. fumigatus clinical isolates from patients failing echinocandin therapy. Echinocandin resistance is known to arise from amino acid substitutions in β-(1,3)-d-glucan synthase encoded by the fks1 gene. Yet these clinical isolates did not contain mutations in fks1, indicating an undefined resistance mechanism. To explore this new mechanism, we used a laboratory-derived strain, RG101, with a nearly identical caspofungin (CAS) susceptibility phenotype that also does not contain fks1 mutations. Glucan synthase isolated from RG101 was fully sensitive to echinocandins. Yet exposure of RG101 to CAS during growth yielded a modified enzyme that was drug insensitive (4 log orders) in kinetic inhibition assays, and this insensitivity was also observed for enzymes isolated from clinical isolates. To understand this alteration, we analyzed whole-enzyme posttranslational modifications (PTMs) but found none linked to resistance. However, analysis of the lipid microenvironment of the enzyme with resistance induced by CAS revealed a prominent increase in the abundances of dihydrosphingosine (DhSph) and phytosphingosine (PhSph). Exogenous addition of DhSph and PhSph to the sensitive enzyme recapitulated the drug insensitivity of the CAS-derived enzyme. Further analysis demonstrated that CAS induces mitochondrion-derived reactive oxygen species (ROS) and that dampening ROS formation by antimycin A or thiourea eliminated drug-induced resistance. We conclude that CAS induces cellular stress, promoting formation of ROS and triggering an alteration in the composition of plasma membrane lipids surrounding glucan synthase, rendering it insensitive to echinocandins.IMPORTANCE Resistance to first-line triazole antifungal agents among Aspergillus species has prompted the use of second-line therapy with echinocandins. As the number of Aspergillus-infected patients treated with echinocandins is rising, clinical observations of drug resistance are also increasing, indicating an emerging global health threat. Our knowledge regarding the development of clinical echinocandin resistance is largely derived from Candida spp., while little is known about resistance in Aspergillus. Therefore, it is important to understand the specific cellular responses raised by A. fumigatus against echinocandins. We discovered a new mechanism of resistance in A. fumigatus that is independent of the well-characterized FKS mutation mechanism observed in Candida This study identified an off-target effect of CAS, i.e., ROS production, and integrated oxidative stress and sphingolipid alterations into a novel mechanism of resistance. This stress-induced response has implications for drug resistance and/or tolerance mechanisms in other fungal pathogens.
Keywords: Aspergillus fumigatus; ROS; antifungal resistance; caspofungin; echinocandins; glucan synthase; glucan synthase inhibitors; lipids.
Copyright © 2019 Satish et al.
Figures
Similar articles
-
Resistance in human pathogenic yeasts and filamentous fungi: prevalence, underlying molecular mechanisms and link to the use of antifungals in humans and the environment.Dan Med J. 2016 Oct;63(10):B5288. Dan Med J. 2016. PMID: 27697142 Review.
-
The Paradoxical Effect of Echinocandins in Aspergillus fumigatus Relies on Recovery of the β-1,3-Glucan Synthase Fks1.Antimicrob Agents Chemother. 2017 Jan 24;61(2):e01690-16. doi: 10.1128/AAC.01690-16. Print 2017 Feb. Antimicrob Agents Chemother. 2017. PMID: 27872079 Free PMC article.
-
Characterization of Aspergillus fumigatus mutants with reduced susceptibility to caspofungin.Med Mycol. 2005 May;43 Suppl 1:S299-305. doi: 10.1080/13693780400029023. Med Mycol. 2005. PMID: 16110824 Review.
-
Emergence of Echinocandin Resistance Due to a Point Mutation in the fks1 Gene of Aspergillus fumigatus in a Patient with Chronic Pulmonary Aspergillosis.Antimicrob Agents Chemother. 2017 Nov 22;61(12):e01277-17. doi: 10.1128/AAC.01277-17. Print 2017 Dec. Antimicrob Agents Chemother. 2017. PMID: 28923871 Free PMC article.
-
Target enzyme mutations confer differential echinocandin susceptibilities in Candida kefyr.Antimicrob Agents Chemother. 2014 Sep;58(9):5421-7. doi: 10.1128/AAC.00096-14. Epub 2014 Jun 30. Antimicrob Agents Chemother. 2014. PMID: 24982083 Free PMC article.
Cited by
-
Genomic and Molecular Identification of Genes Contributing to the Caspofungin Paradoxical Effect in Aspergillus fumigatus.Microbiol Spectr. 2022 Oct 26;10(5):e0051922. doi: 10.1128/spectrum.00519-22. Epub 2022 Sep 12. Microbiol Spectr. 2022. PMID: 36094204 Free PMC article.
-
Aspergillus fumigatus Transcription Factors Involved in the Caspofungin Paradoxical Effect.mBio. 2020 Jun 16;11(3):e00816-20. doi: 10.1128/mBio.00816-20. mBio. 2020. PMID: 32546620 Free PMC article.
-
Novel and potent antimicrobial effects of caspofungin on drug-resistant Candida and bacteria.Sci Rep. 2020 Oct 20;10(1):17745. doi: 10.1038/s41598-020-74749-8. Sci Rep. 2020. PMID: 33082485 Free PMC article.
-
Adaptative survival of Aspergillus fumigatus to echinocandins arises from cell wall remodeling beyond β-1,3-glucan synthesis inhibition.Nat Commun. 2024 Jul 31;15(1):6382. doi: 10.1038/s41467-024-50799-8. Nat Commun. 2024. PMID: 39085213 Free PMC article.
-
FKS1 mutation associated with decreased echinocandin susceptibility of Aspergillus fumigatus following anidulafungin exposure.Sci Rep. 2020 Jul 20;10(1):11976. doi: 10.1038/s41598-020-68706-8. Sci Rep. 2020. PMID: 32686741 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
