

Proteolytic and Opportunistic Breaching of the Basement Membrane Zone by Immune Cells during Tumor Initiation
- PMID: 31167131
- PMCID: PMC6581915
- DOI: 10.1016/j.celrep.2019.05.029
Proteolytic and Opportunistic Breaching of the Basement Membrane Zone by Immune Cells during Tumor Initiation
Abstract
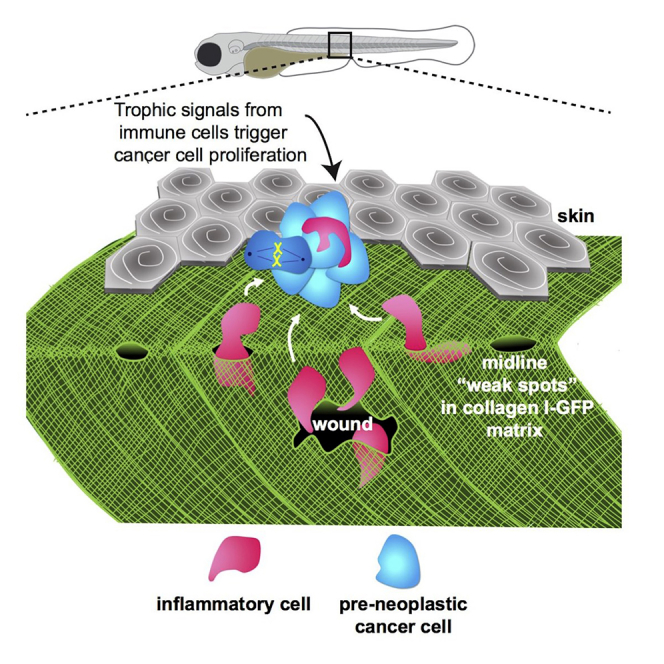
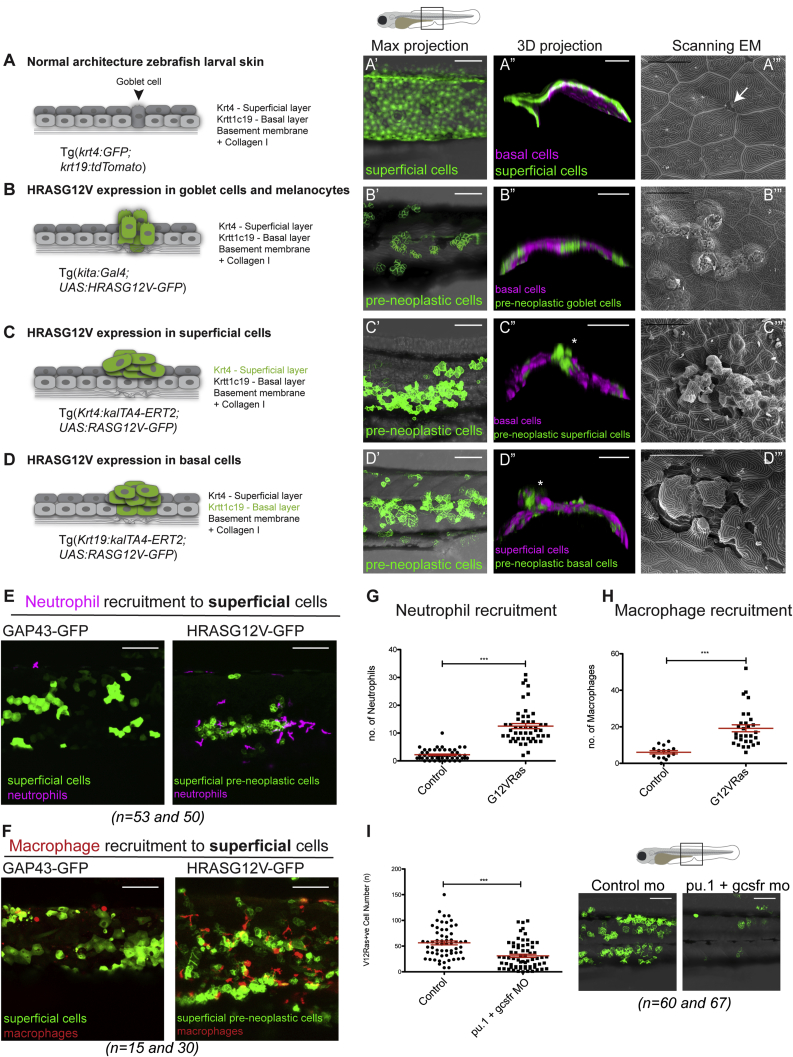
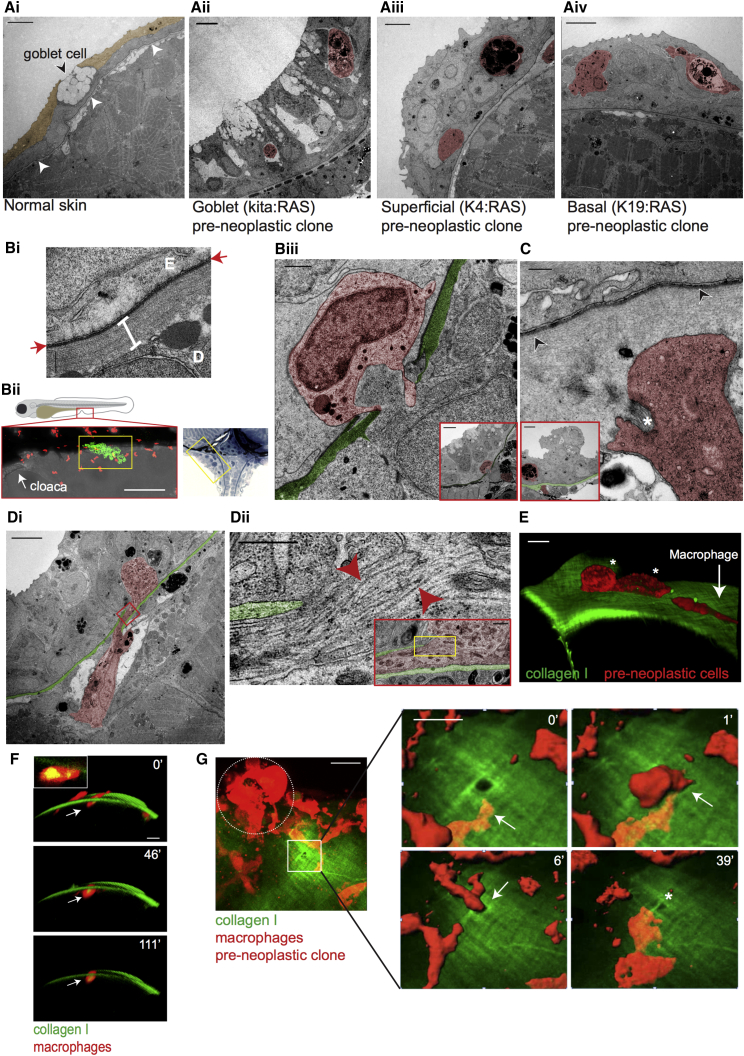
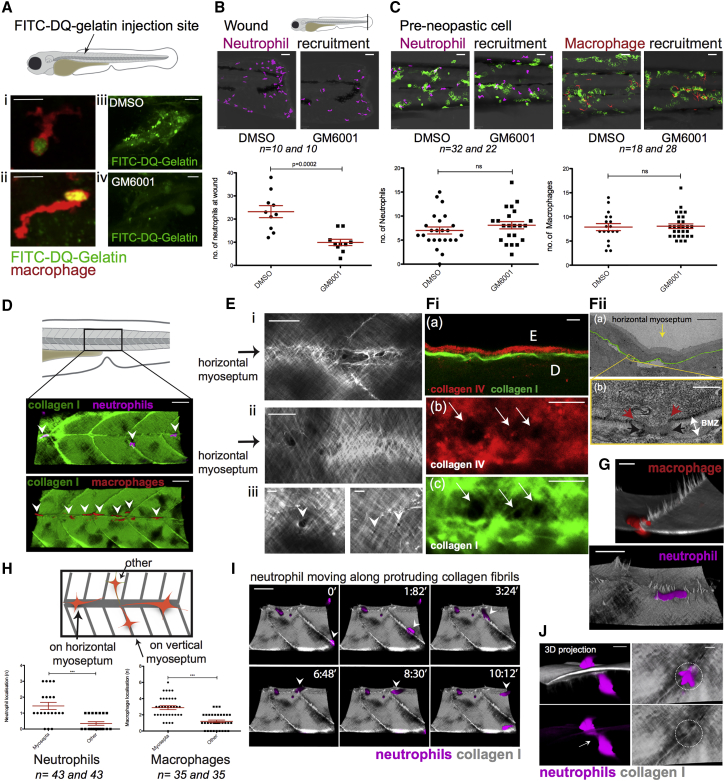
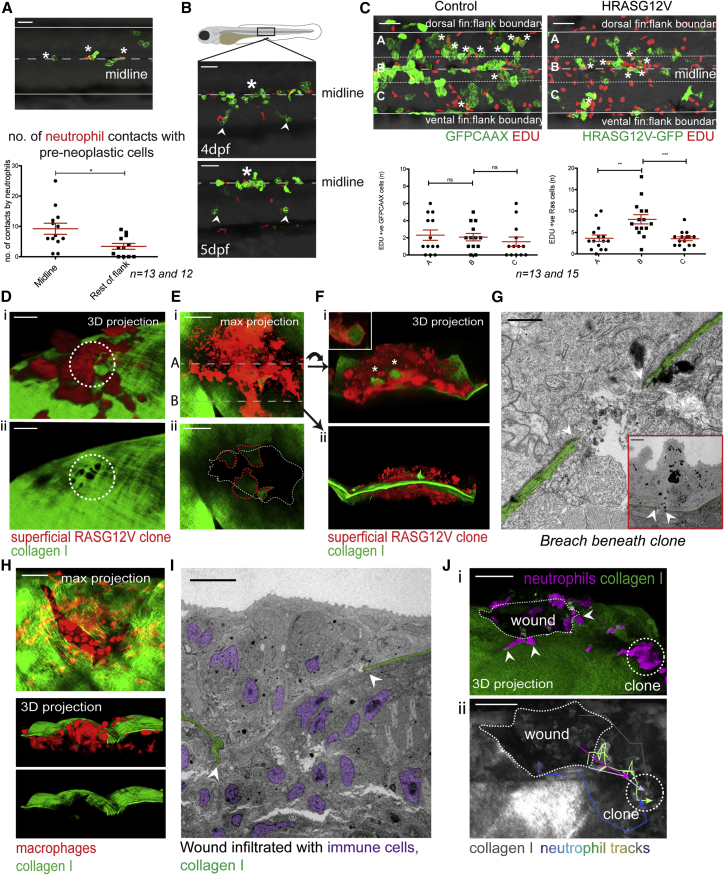
Cancer-related inflammation impacts significantly on cancer development and progression. From early stages, neutrophils and macrophages are drawn to pre-neoplastic cells in the epidermis, but before directly interacting, they must first breach the underlying extracellular matrix barrier layer that includes the basement membrane. Using several different skin cancer models and a collagen I-GFP transgenic zebrafish line, we have undertaken correlative light and electron microscopy (CLEM) to capture the moments when immune cells traverse the basement membrane. We show evidence both for active proteolytic burrowing and for the opportunistic use of pre-existing weak spots in the matrix layer. We show that these small holes, as well as much larger, cancer cell-generated or wound-triggered gaps in the matrix barrier, provide portals for immune cells to access cancer cells in the epidermis and thus are rate limiting in cancer progression.
Keywords: CLEM; basement membrane zone; cancer; cell motility; collagen; inflammation; macrophages; neutrophils; zebrafish.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Antonio N., Bønnelykke-Behrndtz M.L., Ward L.C., Collin J., Christensen I.J., Steiniche T., Schmidt H., Feng Y., Martin P. The wound inflammatory response exacerbates growth of pre-neoplastic cells and progression to cancer. EMBO J. 2015;34:2219–2236. - PMC - PubMed
- Antonio, N., Bonnelykke-Behrndtz, M.L., Ward, L.C., Collin, J., Christensen, I.J., Steiniche, T., Schmidt, H., Feng, Y., and Martin, P. (2015). The wound inflammatory response exacerbates growth of pre-neoplastic cells and progression to cancer. EMBO J. 34, 2219-2236. - PMC - PubMed
-
- Chia K., Mazzolini J., Mione M., Sieger D. Tumor initiating cells induce Cxcr4-mediated infiltration of pro-tumoral macrophages into the brain. eLife. 2018;7 e31918. - PMC - PubMed
- Chia, K., Mazzolini, J., Mione, M., and Sieger, D. (2018). Tumor initiating cells induce Cxcr4-mediated infiltration of pro-tumoral macrophages into the brain. eLife 7, e31918. - PMC - PubMed
-
- Coffelt S.B., Kersten K., Doornebal C.W., Weiden J., Vrijland K., Hau C.S., Verstegen N.J.M., Ciampricotti M., Hawinkels L.J.A.C., Jonkers J., de Visser K.E. IL-17-producing γδ T cells and neutrophils conspire to promote breast cancer metastasis. Nature. 2015;522:345–348. - PMC - PubMed
- Coffelt, S.B., Kersten, K., Doornebal, C.W., Weiden, J., Vrijland, K., Hau, C.S., Verstegen, N.J.M., Ciampricotti, M., Hawinkels, L.J.A.C., Jonkers, J., and de Visser, K.E. (2015). IL-17-producing γδ T cells and neutrophils conspire to promote breast cancer metastasis. Nature 522, 345-348. - PMC - PubMed
-
- Distel M., Wullimann M.F., Koster R.W. Optimized Gal4 genetics for permanent gene expression mapping in zebrafish. PNAS. 2009 Aug 11;106(32):13365–13370. - PMC - PubMed
- Distel, M., Wullimann, MF., Koster RW. Optimized Gal4 genetics for permanent gene expression mapping in zebrafish. 2009. PNAS Aug 11;106(32):13365-70. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
