

Simvastatin Attenuates Cardiac Fibrosis via Regulation of Cardiomyocyte-Derived Exosome Secretion
- PMID: 31167519
- PMCID: PMC6617127
- DOI: 10.3390/jcm8060794
Simvastatin Attenuates Cardiac Fibrosis via Regulation of Cardiomyocyte-Derived Exosome Secretion
Abstract
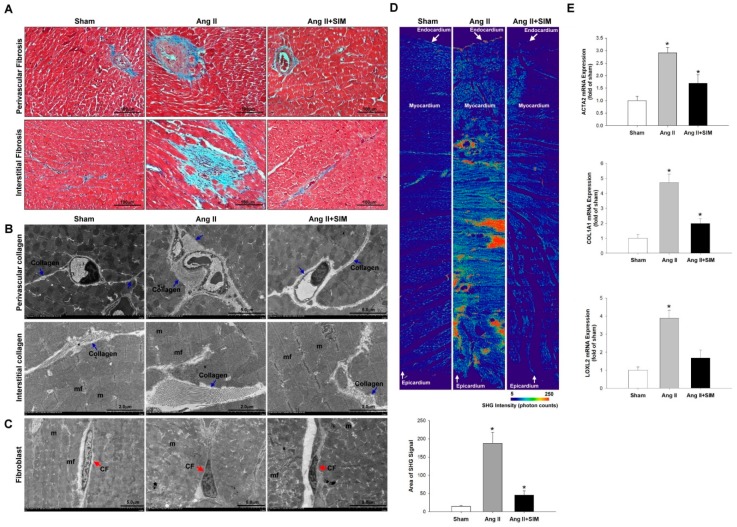
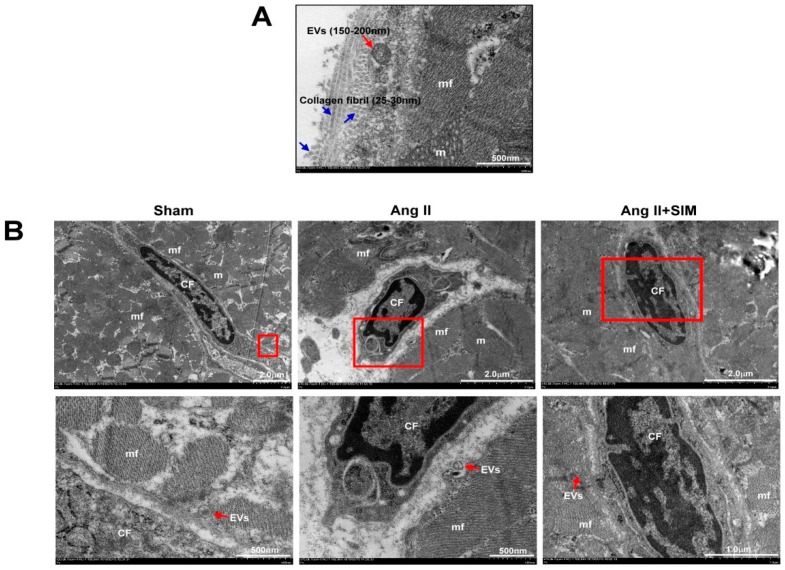
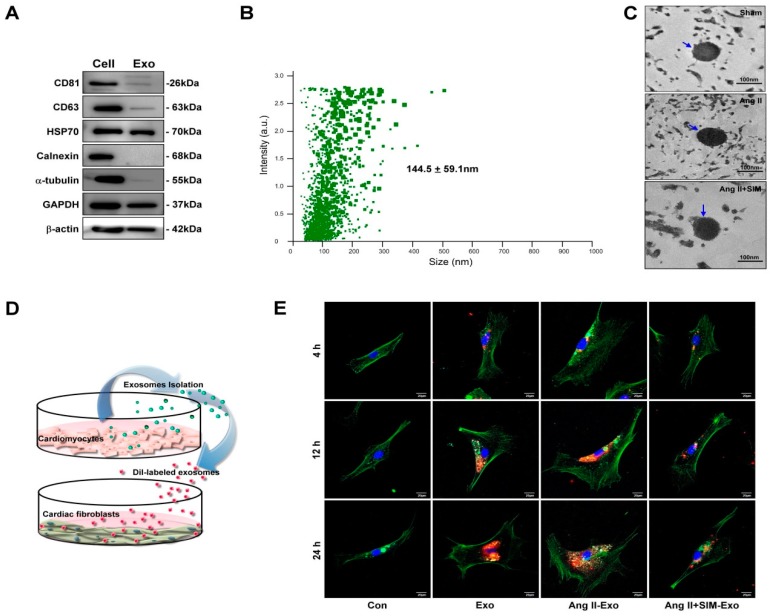
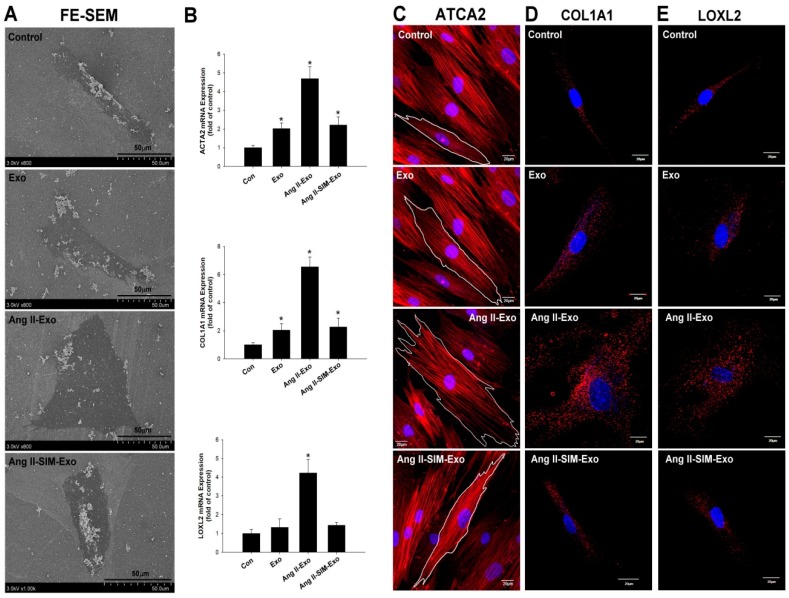
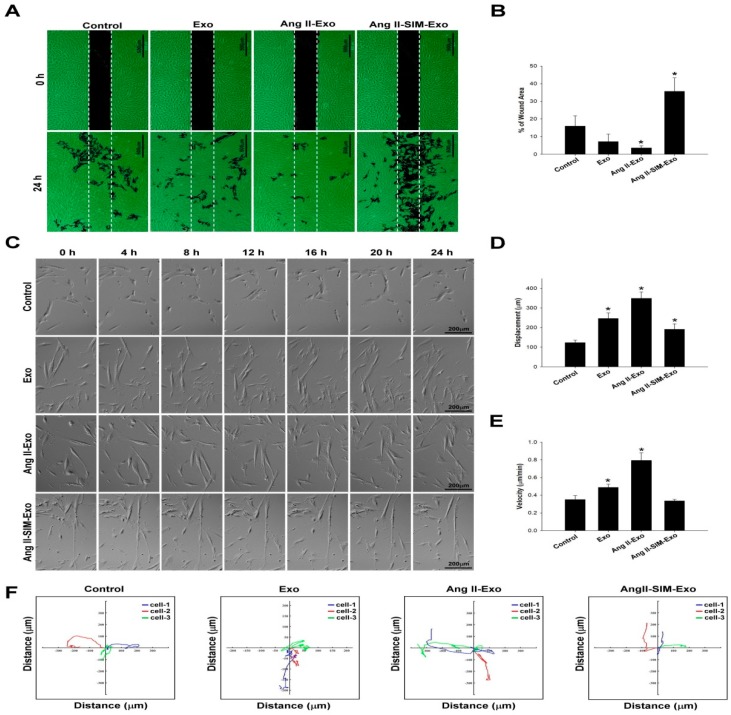
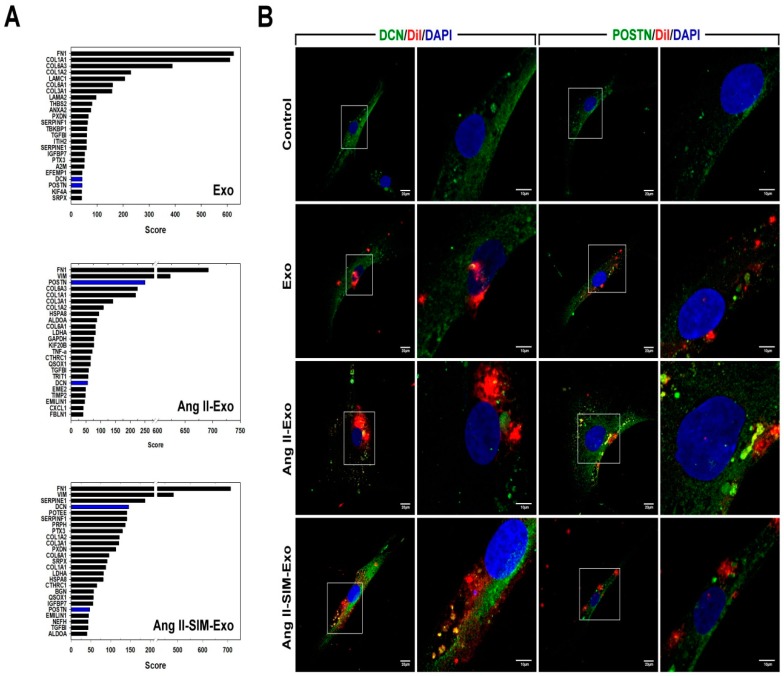
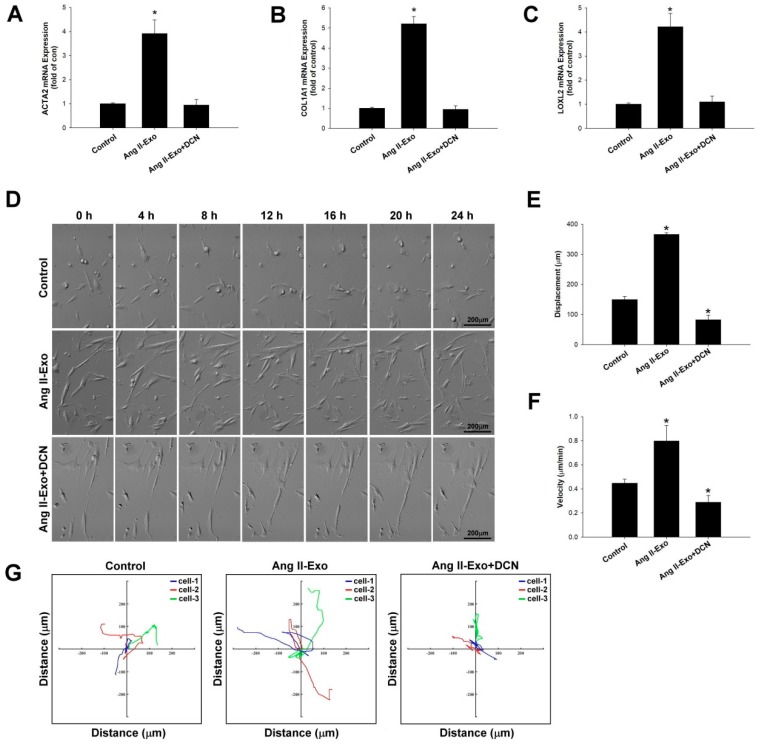
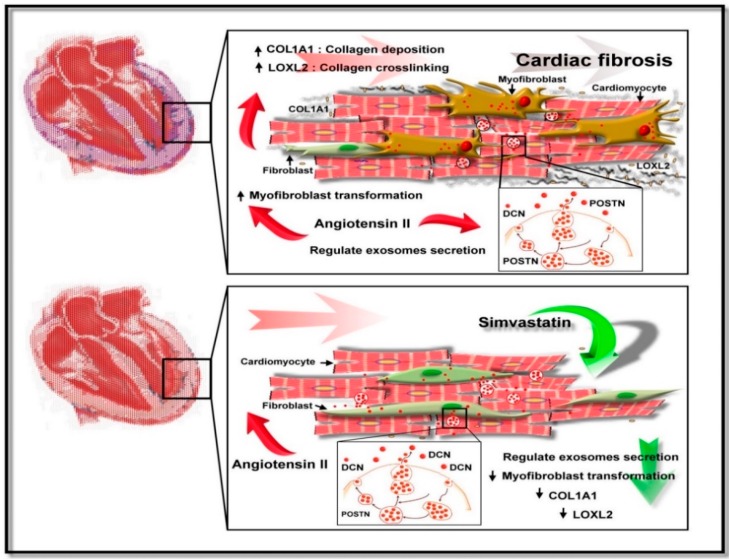
Exosome-mediated communication within the cardiac microenvironment is associated with cardiac fibrosis. Simvastatin (SIM), a potent statin, protects against cardiac fibrosis, but its mechanism of action is unclear. We investigated the inhibitory effects and underlying mechanism of simvastatin in cardiac fibrosis, by regulating exosome-mediated communication. Male Sprague-Dawley rats were treated with angiotensin (Ang) II alone, or with SIM for 28 d. Cardiac fibrosis, expressions of fibrosis-associated proteins and mRNAs, and collagen fiber arrangement and deposition were examined. Protein expressions in exosomes isolated from Ang II-treated cardiomyocytes (CMs) were evaluated using nano-ultra-performance liquid chromatographic system, combined with tandem mass spectrometry. Transformation of fibroblasts to myofibroblasts was evaluated using scanning electron and confocal microscopy, and migration assays. Our results showed that SIM attenuated in vivo expression of collagen and collagen-associated protein, as well as collagen deposition, and cardiac fibrosis. The statin also upregulated decorin and downregulated periostin in CM-derived exosomes. Furthermore, it suppressed Ang II-induced transformation of fibroblast to myofibroblast, as well as fibroblast migration. Exosome-mediated cell-cell communication within the cardiac tissue critically regulated cardiac fibrosis. Specifically, SIM regulated the release of CM exosomes, and attenuated Ang II-induced cardiac fibrosis, highlighting its potential as a novel therapy for cardiac fibrosis.
Keywords: cardiac fibrosis; decorin; exosomes; periostin; statins.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
Grants and funding
- MOST104-2314-B-037-087/Ministry of Science and Technology, Taiwan
- MOST105-2314-B-037-046/Ministry of Science and Technology, Taiwan
- MOST104-2314-B-037-081-MY2/Ministry of Science and Technology, Taiwan
- KMUH102-2M18/Kaohsiung Medical University Chung-Ho Memorial Hospital
- KMTTH100-018/Kaohsiung Municipal Ta-Tung Hospital Research Foundation
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
