

Tau is required for progressive synaptic and memory deficits in a transgenic mouse model of α-synucleinopathy
- PMID: 31168644
- PMCID: PMC6778173
- DOI: 10.1007/s00401-019-02032-w
Tau is required for progressive synaptic and memory deficits in a transgenic mouse model of α-synucleinopathy
Abstract
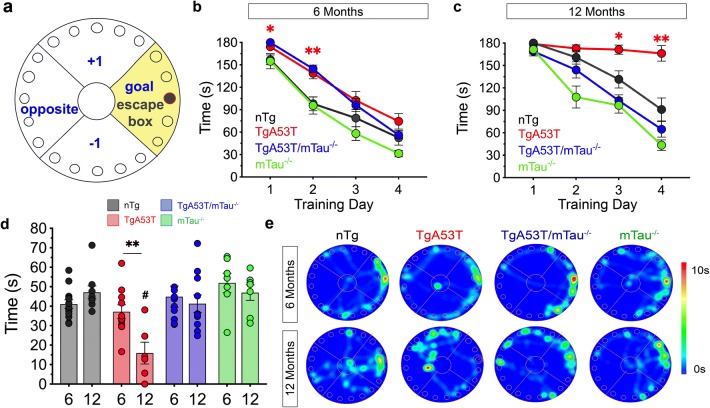
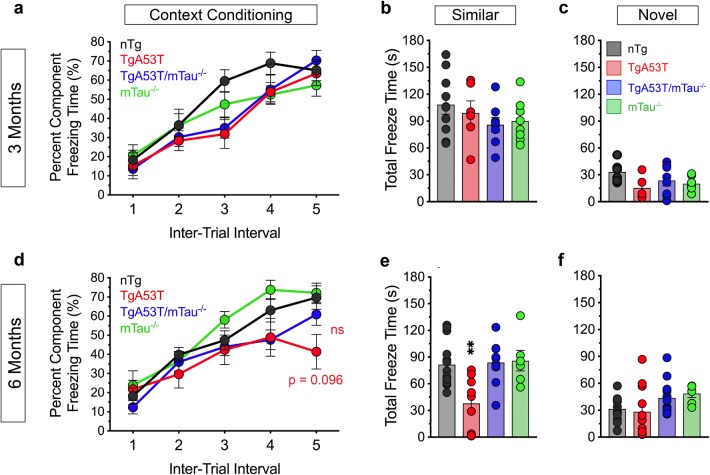
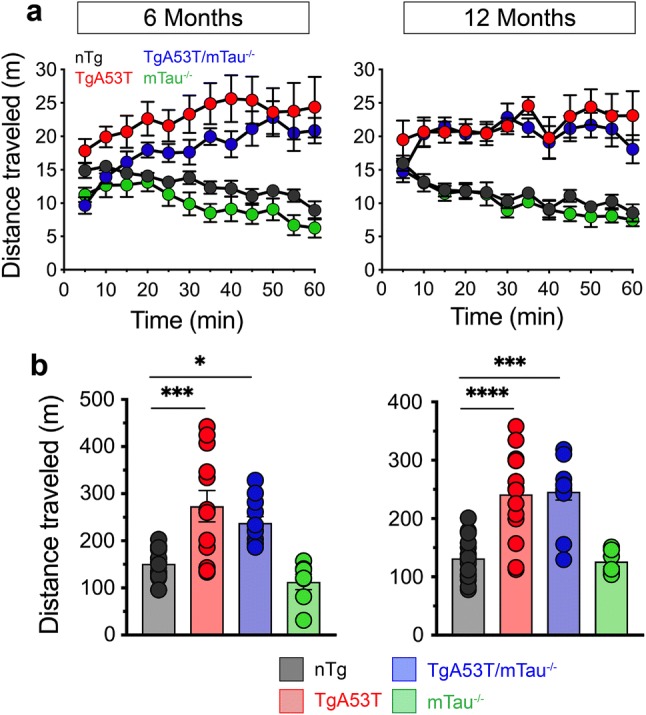
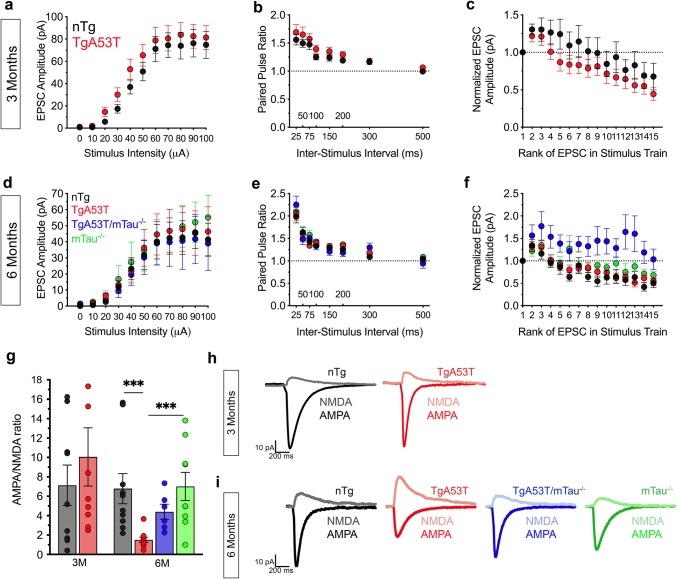
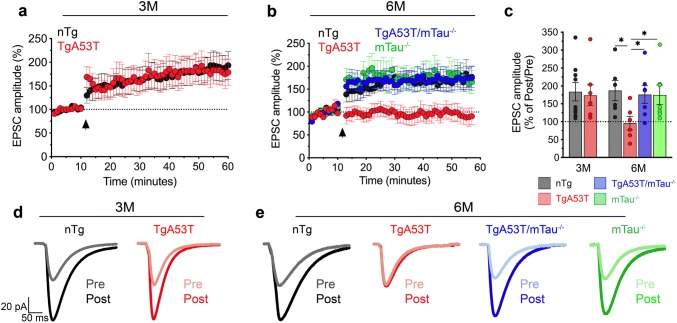
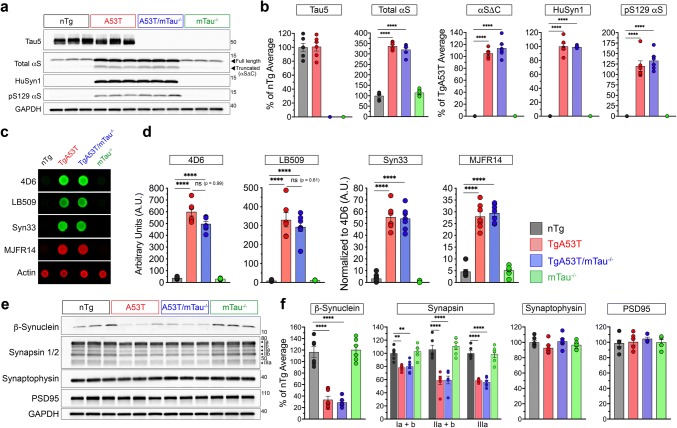
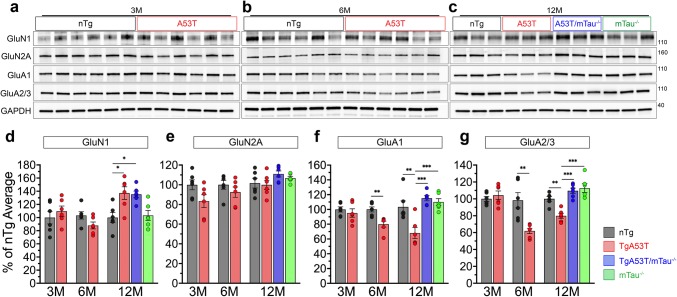
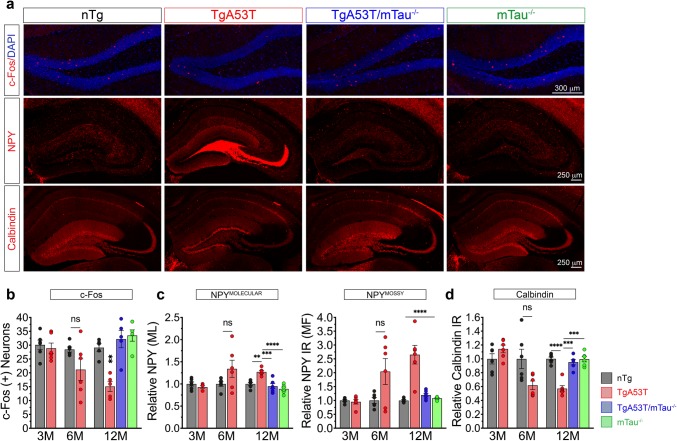
Parkinson's disease dementia (PDD) and dementia with Lewy bodies (DLB) are clinically and neuropathologically highly related α-synucleinopathies that collectively constitute the second leading cause of neurodegenerative dementias. Genetic and neuropathological studies directly implicate α-synuclein (αS) abnormalities in PDD and DLB pathogenesis. However, it is currently unknown how αS abnormalities contribute to memory loss, particularly since forebrain neuronal loss in PDD and DLB is less severe than in Alzheimer's disease. Previously, we found that familial Parkinson's disease-linked human mutant A53T αS causes aberrant localization of the microtubule-associated protein tau to postsynaptic spines in neurons, leading to postsynaptic deficits. Thus, we directly tested if the synaptic and memory deficits in a mouse model of α-synucleinopathy (TgA53T) are mediated by tau. TgA53T mice exhibit progressive memory deficits associated with postsynaptic deficits in the absence of obvious neuropathological and neurodegenerative changes in the hippocampus. Significantly, removal of endogenous mouse tau expression in TgA53T mice (TgA53T/mTau-/-), achieved by mating TgA53T mice to mouse tau-knockout mice, completely ameliorates cognitive dysfunction and concurrent synaptic deficits without affecting αS expression or accumulation of selected toxic αS oligomers. Among the known tau-dependent effects, memory deficits in TgA53T mice were associated with hippocampal circuit remodeling linked to chronic network hyperexcitability. This remodeling was absent in TgA53T/mTau-/- mice, indicating that postsynaptic deficits, aberrant network hyperactivity, and memory deficits are mechanistically linked. Our results directly implicate tau as a mediator of specific human mutant A53T αS-mediated abnormalities related to deficits in hippocampal neurotransmission and suggest a mechanism for memory impairment that occurs as a consequence of synaptic dysfunction rather than synaptic or neuronal loss. We hypothesize that these initial synaptic deficits contribute to network hyperexcitability which, in turn, exacerbate cognitive dysfunction. Our results indicate that these synaptic changes present potential therapeutic targets for amelioration of memory deficits in α-synucleinopathies.
Keywords: Dementia; Lewy body disease; Neuronal plasticity; Parkinson’s disease; Tau; α-Synuclein.
Conflict of interest statement
SEL is a scientific consultant for Acelot, Inc. and has no actual or potential conflict of interest in relation to this study. All other authors declare no conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
