

Proteasome-dependent degradation of intracellular carbamylated proteins
- PMID: 31170093
- PMCID: PMC6594819
- DOI: 10.18632/aging.102002
Proteasome-dependent degradation of intracellular carbamylated proteins
Abstract
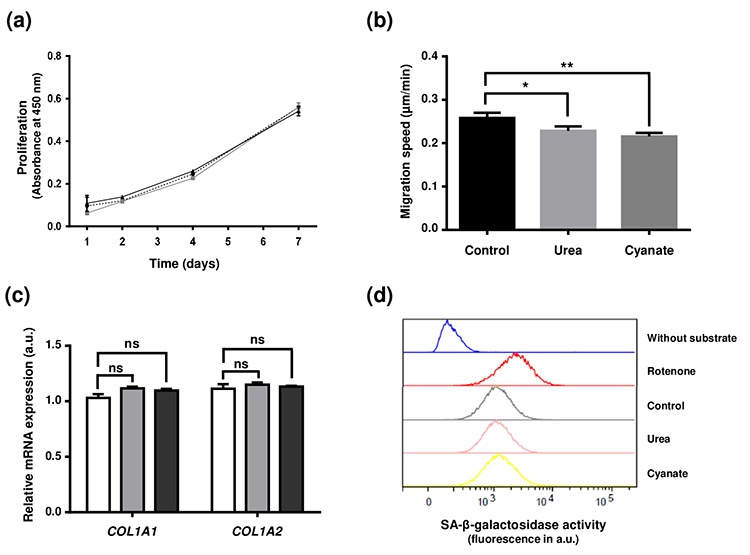
Carbamylation, which corresponds to the binding of isocyanic acid to the amino groups of proteins, is a nonenzymatic post-translational modification responsible for alterations of protein structural and functional properties. Tissue accumulation of carbamylation-derived products and their role in pathological processes such as atherosclerosis or chronic renal failure have been previously documented. However, few studies have focused on the carbamylation of intracellular proteins and their subsequent role in cellular aging. This study aimed to determine the extent of intracellular protein carbamylation, its impact on cell functions and the ability of cells to degrade these modified proteins. Fibroblasts were incubated with cyanate or urea and the carbamylation level was evaluated by immunostaining and homocitrulline quantification. The results showed that carbamylated proteins accumulated intracellularly and that all proteins were susceptible. The presence of intracellular carbamylated proteins did not modify cell proliferation or type I collagen synthesis nor did it induce cell senescence, but it significantly decreased cell motility. Fibroblasts were able to degrade carbamylated proteins through the ubiquitin-proteasome system. In conclusion, intracellular proteins are susceptible to carbamylation but their accumulation does not seem to deeply affect cell function, owing largely to their elimination by the ubiquitin-proteasome system.
Keywords: carbamylation; cell aging; homocitrulline; nonenzymatic post-translational modifications; proteasome; proteostasis.
Conflict of interest statement
Figures






References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
