

Disruption of the NlpD lipoprotein of the plague pathogen Yersinia pestis affects iron acquisition and the activity of the twin-arginine translocation system
- PMID: 31170147
- PMCID: PMC6553720
- DOI: 10.1371/journal.pntd.0007449
Disruption of the NlpD lipoprotein of the plague pathogen Yersinia pestis affects iron acquisition and the activity of the twin-arginine translocation system
Abstract
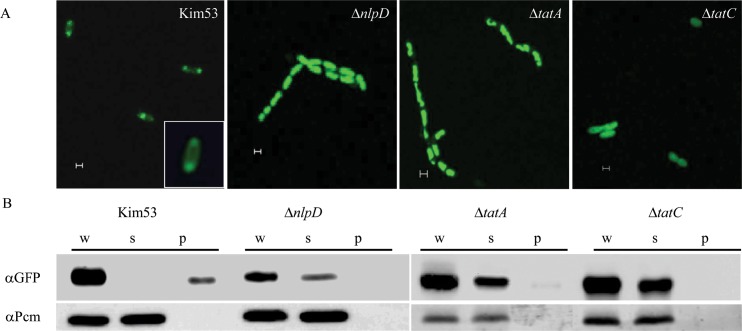
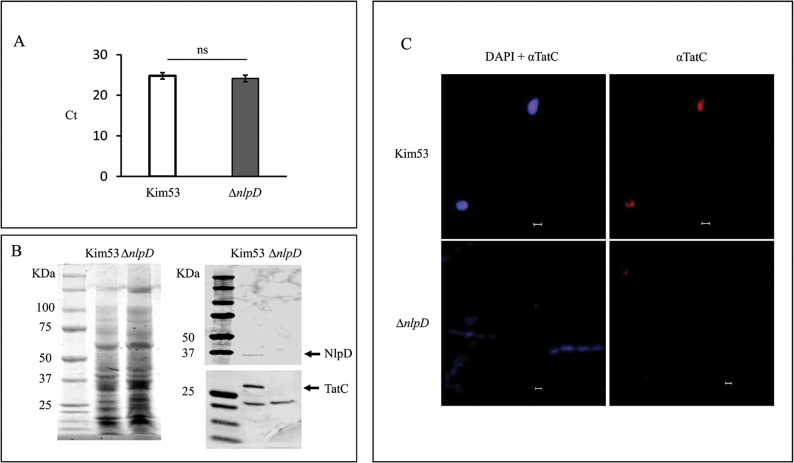
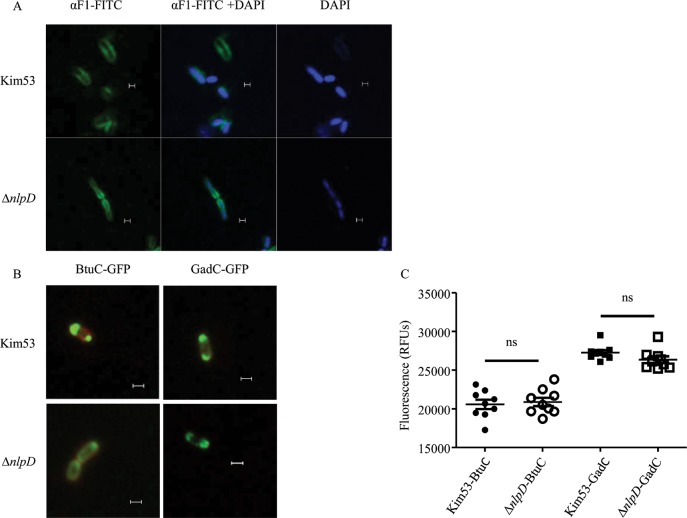
We have previously shown that the cell morphogenesis NlpD lipoprotein is essential for virulence of the plague bacteria, Yersinia pestis. To elucidate the role of NlpD in Y. pestis pathogenicity, we conducted a whole-genome comparative transcriptome analysis of the wild-type Y. pestis strain and an nlpD mutant under conditions mimicking early stages of infection. The analysis suggested that NlpD is involved in three phenomena: (i) Envelope stability/integrity evidenced by compensatory up-regulation of the Cpx and Psp membrane stress-response systems in the mutant; (ii) iron acquisition, supported by modulation of iron metabolism genes and by limited growth in iron-deprived medium; (iii) activity of the twin-arginine (Tat) system, which translocates folded proteins across the cytoplasmic membrane. Virulence studies of Y. pestis strains mutated in individual Tat components clearly indicated that the Tat system is central in Y. pestis pathogenicity and substantiated the assumption that NlpD essentiality in iron utilization involves the activity of the Tat system. This study reveals a new role for NlpD in Tat system activity and iron assimilation suggesting a modality by which this lipoprotein is involved in Y. pestis pathogenesis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
The NlpD lipoprotein is a novel Yersinia pestis virulence factor essential for the development of plague.PLoS One. 2009 Sep 14;4(9):e7023. doi: 10.1371/journal.pone.0007023. PLoS One. 2009. PMID: 19759820 Free PMC article.
-
Manganese transporters Yfe and MntH are Fur-regulated and important for the virulence of Yersinia pestis.Microbiology (Reading). 2012 Mar;158(Pt 3):804-815. doi: 10.1099/mic.0.053710-0. Epub 2012 Jan 5. Microbiology (Reading). 2012. PMID: 22222497 Free PMC article.
-
The Yfe and Feo transporters are involved in microaerobic growth and virulence of Yersinia pestis in bubonic plague.Infect Immun. 2012 Nov;80(11):3880-91. doi: 10.1128/IAI.00086-12. Epub 2012 Aug 27. Infect Immun. 2012. PMID: 22927049 Free PMC article.
-
Yersiniabactin iron uptake: mechanisms and role in Yersinia pestis pathogenesis.Microbes Infect. 2011 Sep;13(10):808-17. doi: 10.1016/j.micinf.2011.04.008. Epub 2011 May 12. Microbes Infect. 2011. PMID: 21609780 Free PMC article. Review.
-
Contributions of Yersinia pestis outer membrane protein Ail to plague pathogenesis.Curr Opin Infect Dis. 2022 Jun 1;35(3):188-195. doi: 10.1097/QCO.0000000000000830. Curr Opin Infect Dis. 2022. PMID: 35665712 Free PMC article. Review.
Cited by
-
Screening of an FDA-Approved Library for Novel Drugs against Y. pestis.Antibiotics (Basel). 2021 Jan 3;10(1):40. doi: 10.3390/antibiotics10010040. Antibiotics (Basel). 2021. PMID: 33401634 Free PMC article.
-
One fold, many functions-M23 family of peptidoglycan hydrolases.Front Microbiol. 2022 Oct 21;13:1036964. doi: 10.3389/fmicb.2022.1036964. eCollection 2022. Front Microbiol. 2022. PMID: 36386627 Free PMC article. Review.
-
Cpx-signalling facilitates Hms-dependent biofilm formation by Yersinia pseudotuberculosis.NPJ Biofilms Microbiomes. 2022 Mar 29;8(1):13. doi: 10.1038/s41522-022-00281-4. NPJ Biofilms Microbiomes. 2022. PMID: 35351893 Free PMC article.
-
Secreted LysM proteins are required for niche competition and full virulence in Pseudomonas savastanoi during host plant infection.PLoS Pathog. 2025 Aug 1;21(8):e1013121. doi: 10.1371/journal.ppat.1013121. eCollection 2025 Aug. PLoS Pathog. 2025. PMID: 40749019 Free PMC article.
-
Searching for putative virulence factors in the genomes of Shewanella indica and Shewanella algae.Arch Microbiol. 2021 Mar;203(2):683-692. doi: 10.1007/s00203-020-02060-1. Epub 2020 Oct 10. Arch Microbiol. 2021. PMID: 33040180
References
-
- Bertherat J (2016) The Challenge of Early Diagnosis of Cushing Disease Recurrence! Endocr Pract 22: 1356–1357. 10.4158/EP161600.CO - DOI - PubMed
-
- Simond P (1905) La question du ve'hicule de la peste. Re Me'dico-Chirurg Bre'sil Am Lat (5).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
