

At the Heart of Bacterial Cytokinesis: The Z Ring
- PMID: 31171437
- PMCID: PMC6831097
- DOI: 10.1016/j.tim.2019.04.011
At the Heart of Bacterial Cytokinesis: The Z Ring
Abstract
Bacterial cell division is mediated by the divisome which is organized by the Z ring, a cytoskeletal element formed by the polymerization of the tubulin homologue FtsZ. Despite billions of years of bacterial evolution, the Z ring is nearly universal among bacteria that have a cell wall and divide by binary fission. Recent studies have revealed the mechanism of cooperative assembly of FtsZ and that the Z ring consists of patches of FtsZ filaments tethered to the membrane that treadmill to distribute the septal biosynthetic machinery. Here, we summarize these advances and discuss questions raised by these new findings.
Keywords: FtsZ; Z ring; treadmilling; tubulin.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Figures
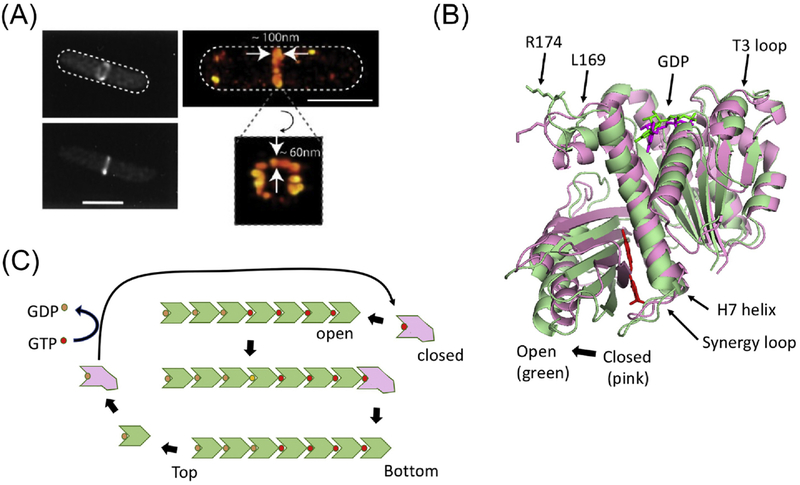
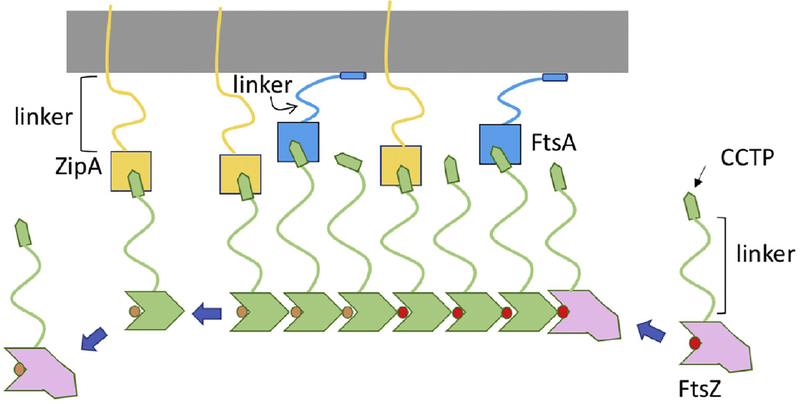
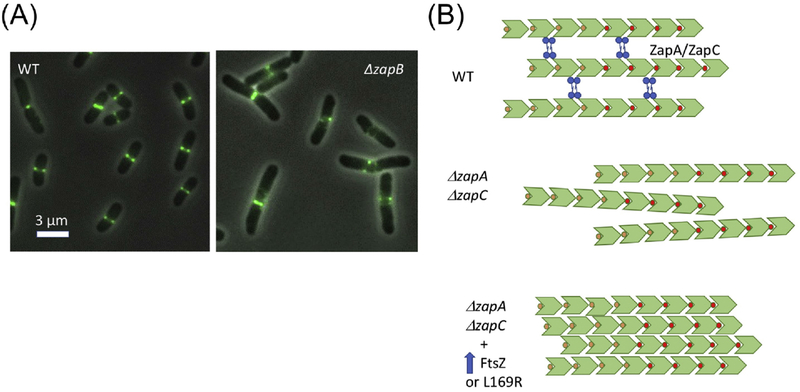
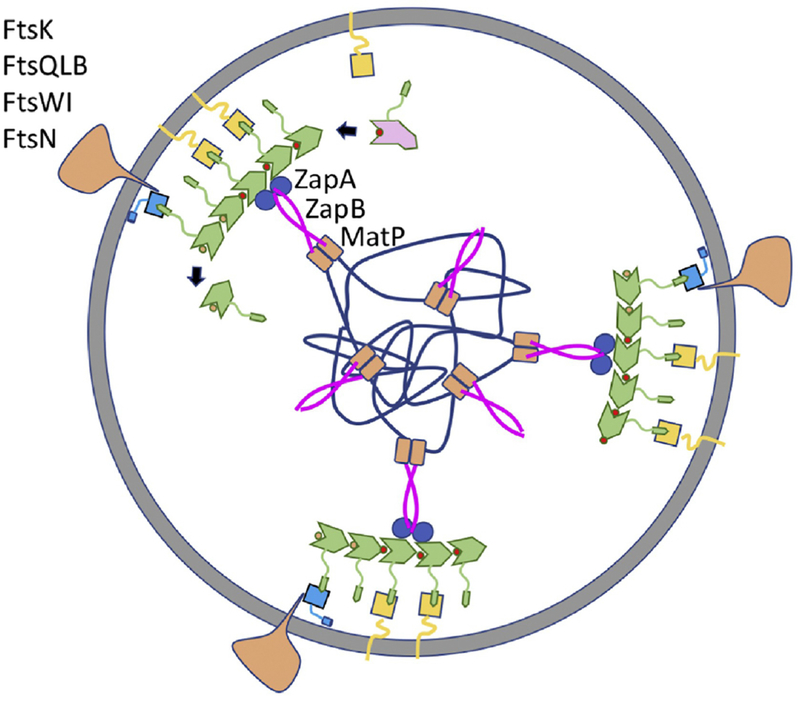
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
