

Editosome RNase III domain interactions are essential for editing and differ between life cycle stages in Trypanosoma brucei
- PMID: 31171708
- PMCID: PMC6800513
- DOI: 10.1261/rna.071258.119
Editosome RNase III domain interactions are essential for editing and differ between life cycle stages in Trypanosoma brucei
Abstract
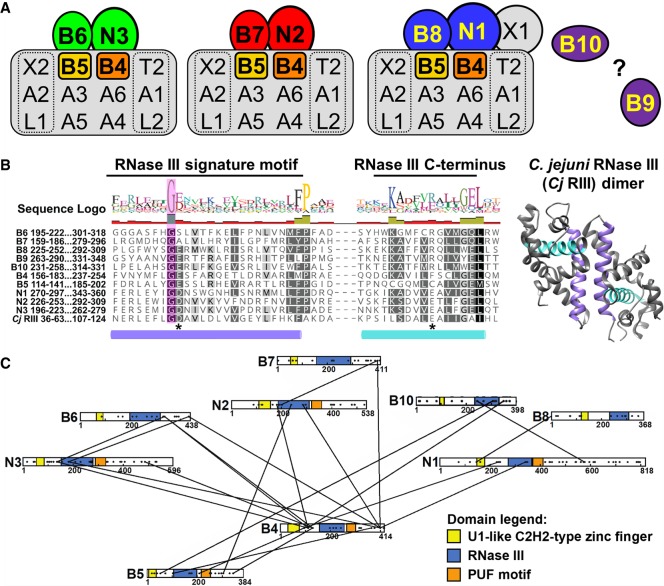
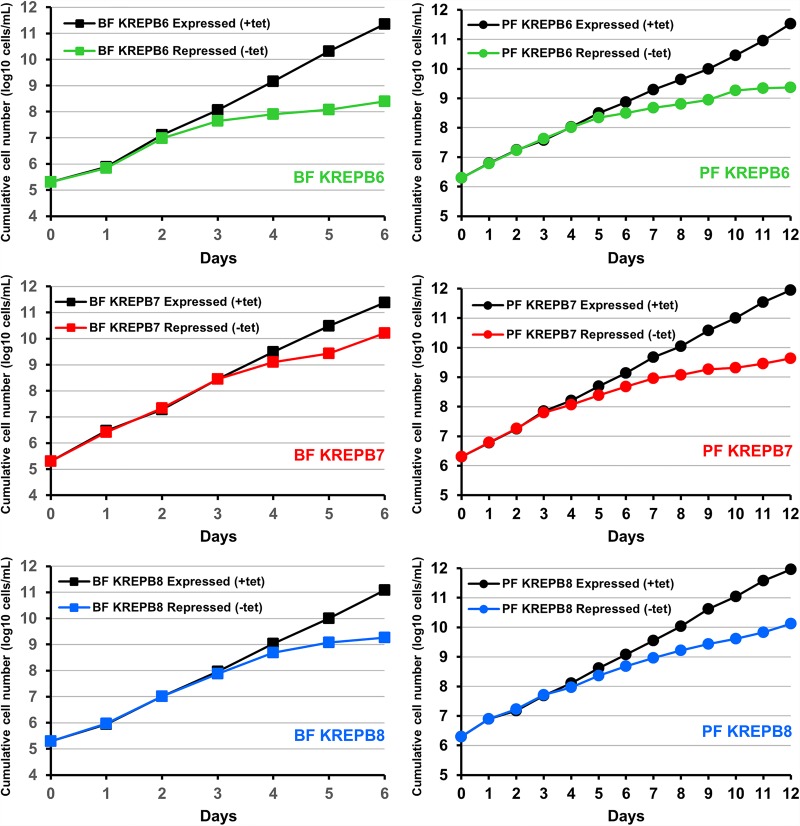
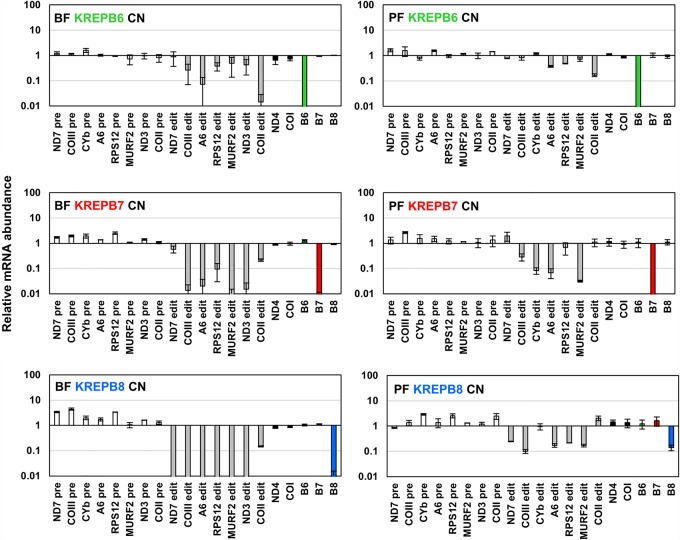
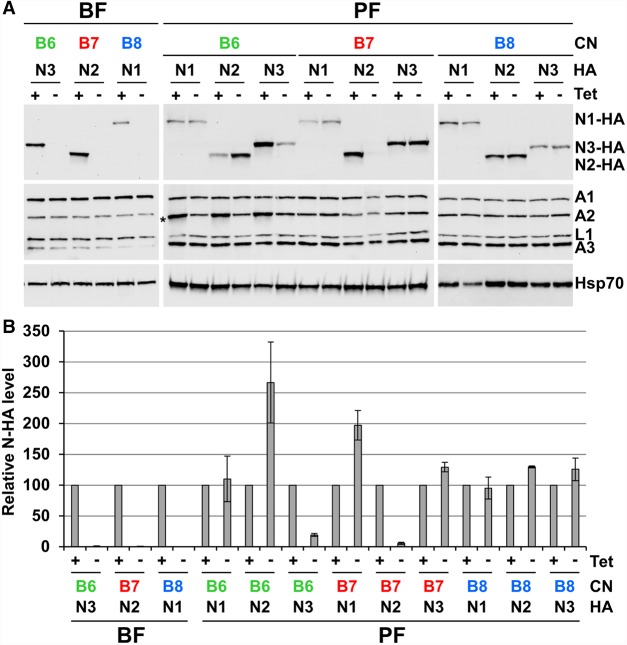
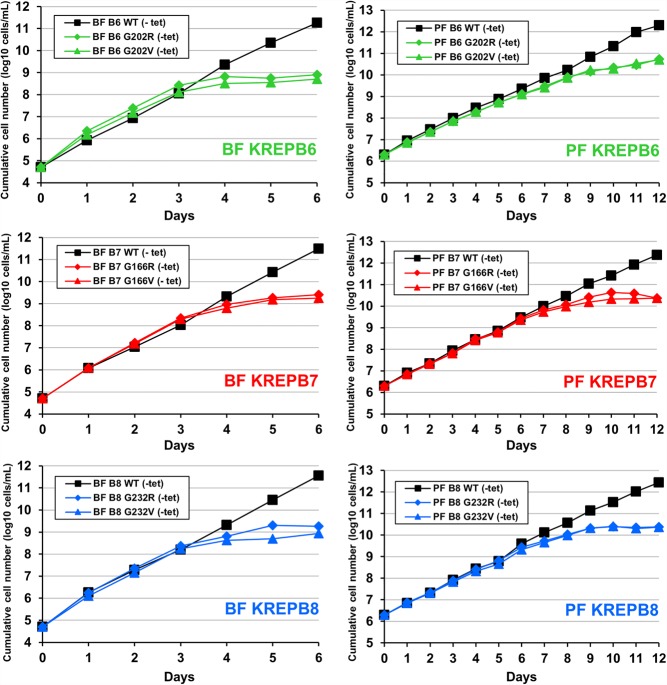
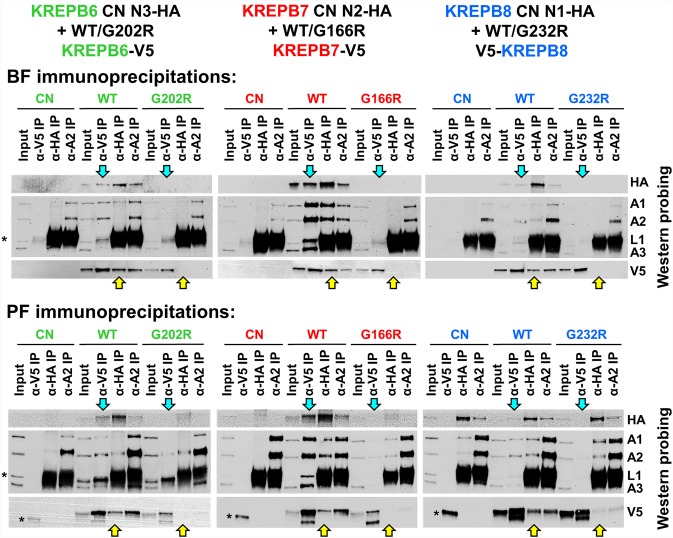
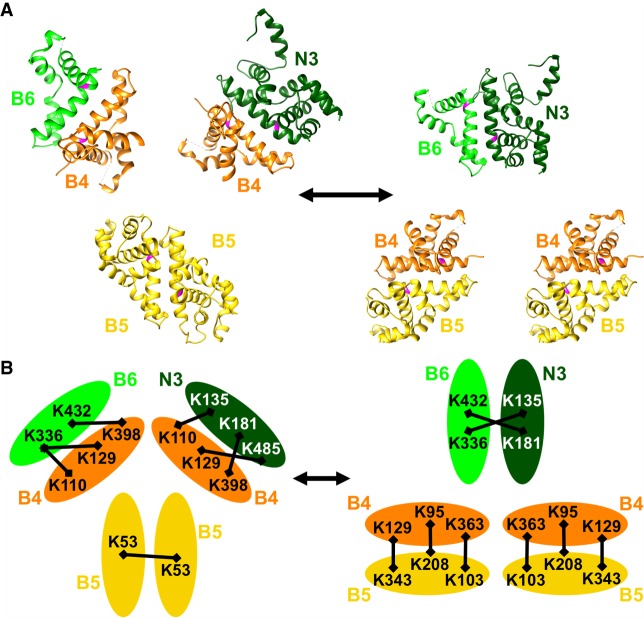
Multiprotein editosomes catalyze gRNA-specified insertion and deletion of uridines to create functional mitochondrial mRNAs in Trypanosoma brucei Three functionally distinct editosomes are distinguished by their single KREN1, KREN2, or KREN3 RNase III endonuclease and, respectively, KREPB8, KREPB7, and KREPB6 partner proteins. These endonucleases perform the first catalytic step of editing, cleaving mRNA in diverse mRNA/gRNA heteroduplex substrates. We identified divergent and likely noncatalytic RNase III domains in KREPB4, KREPB5, KREPB6, KREPB7, KREPB8, KREPB9, and KREPB10 editosome proteins. Because known RNase III endonuclease functional domains are dimeric, the editing endonucleases may form heterodimers with one or more of these divergent RNase III proteins. We show here using conditional null cell lines that KREPB6, KREPB7, and KREPB8 are essential in both procyclic form (PF) and bloodstream (BF) cells. Loss of these proteins results in growth defects and loss of editing in vivo, as does mutation of their RNase III domain that is predicted to prevent dimerization. Loss of KREPB6, KREPB7, or KREPB8 also dramatically reduces cognate endonuclease abundance, as does the RNase III mutation, indicating that RNase III interactions with their partner proteins stabilize the endonucleases. The phenotypic consequences of repression are more severe in BF than in PF, indicating differences in endonuclease function between developmental stages that could impact regulation of editing. These results suggest that KREPB6, KREPB7, and KREPB8 form heterodimers with their respective endonucleases to perform mRNA cleavage. We also present a model wherein editosome proteins with divergent RNase III domains function in substrate selection via enzyme-pseudoenzyme interactions.
Keywords: RNA editing; RNase III; developmental regulation; endonuclease; mitochondria; trypanosomes.
© 2019 McDermott et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
Similar articles
-
KREPB6, KREPB7, and KREPB8 are important for editing endonuclease function in Trypanosoma brucei.RNA. 2012 Feb;18(2):308-20. doi: 10.1261/rna.029314.111. Epub 2011 Dec 19. RNA. 2012. PMID: 22184461 Free PMC article.
-
Endonuclease associations with three distinct editosomes in Trypanosoma brucei.J Biol Chem. 2011 Jun 3;286(22):19320-30. doi: 10.1074/jbc.M111.228965. Epub 2011 Apr 7. J Biol Chem. 2011. PMID: 21474442 Free PMC article.
-
The essential functions of KREPB4 are developmentally distinct and required for endonuclease association with editosomes.RNA. 2017 Nov;23(11):1672-1684. doi: 10.1261/rna.062786.117. Epub 2017 Aug 11. RNA. 2017. PMID: 28802260 Free PMC article.
-
Mitochondrial RNA editing in trypanosomes: small RNAs in control.Biochimie. 2014 May;100:125-31. doi: 10.1016/j.biochi.2014.01.003. Epub 2014 Jan 17. Biochimie. 2014. PMID: 24440637 Free PMC article. Review.
-
Uridine insertion/deletion editing in trypanosomes: a playground for RNA-guided information transfer.Wiley Interdiscip Rev RNA. 2011 Sep-Oct;2(5):669-85. doi: 10.1002/wrna.82. Epub 2011 Mar 23. Wiley Interdiscip Rev RNA. 2011. PMID: 21823228 Free PMC article. Review.
Cited by
-
Domain function and predicted structure of three heterodimeric endonuclease subunits of RNA editing catalytic complexes in Trypanosoma brucei.Nucleic Acids Res. 2022 Sep 23;50(17):10123-10139. doi: 10.1093/nar/gkac753. Nucleic Acids Res. 2022. PMID: 36095119 Free PMC article.
-
Developmental regulation of edited CYb and COIII mitochondrial mRNAs is achieved by distinct mechanisms in Trypanosoma brucei.Nucleic Acids Res. 2020 Sep 4;48(15):8704-8723. doi: 10.1093/nar/gkaa641. Nucleic Acids Res. 2020. PMID: 32738044 Free PMC article.
-
Deep mutational scanning of the RNase III-like domain in Trypanosoma brucei RNA editing protein KREPB4.Front Cell Infect Microbiol. 2024 Apr 8;14:1381155. doi: 10.3389/fcimb.2024.1381155. eCollection 2024. Front Cell Infect Microbiol. 2024. PMID: 38650737 Free PMC article.
-
KREH2 helicase represses ND7 mRNA editing in procyclic-stage Trypanosoma brucei by opposite modulation of canonical and 'moonlighting' gRNA utilization creating a proposed mRNA structure.Nucleic Acids Res. 2024 Oct 28;52(19):11940-11959. doi: 10.1093/nar/gkae699. Nucleic Acids Res. 2024. PMID: 39149912 Free PMC article.
-
Structural Basis of Protein Arginine Methyltransferase Activation by a Catalytically Dead Homolog (Prozyme).J Mol Biol. 2020 Jan 17;432(2):410-426. doi: 10.1016/j.jmb.2019.11.002. Epub 2019 Nov 11. J Mol Biol. 2020. PMID: 31726063 Free PMC article.
References
-
- Brun R, Schönenberger. 1979. Cultivation and in vitro cloning or procyclic culture forms of Trypanosoma brucei in a semi-defined medium. Short communication. Acta Trop 36: 289–292. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous