

The alteration of RhoA geranylgeranylation and Ras farnesylation breaks the integrity of the blood-testis barrier and results in hypospermatogenesis
- PMID: 31171774
- PMCID: PMC6554403
- DOI: 10.1038/s41419-019-1688-9
The alteration of RhoA geranylgeranylation and Ras farnesylation breaks the integrity of the blood-testis barrier and results in hypospermatogenesis
Abstract
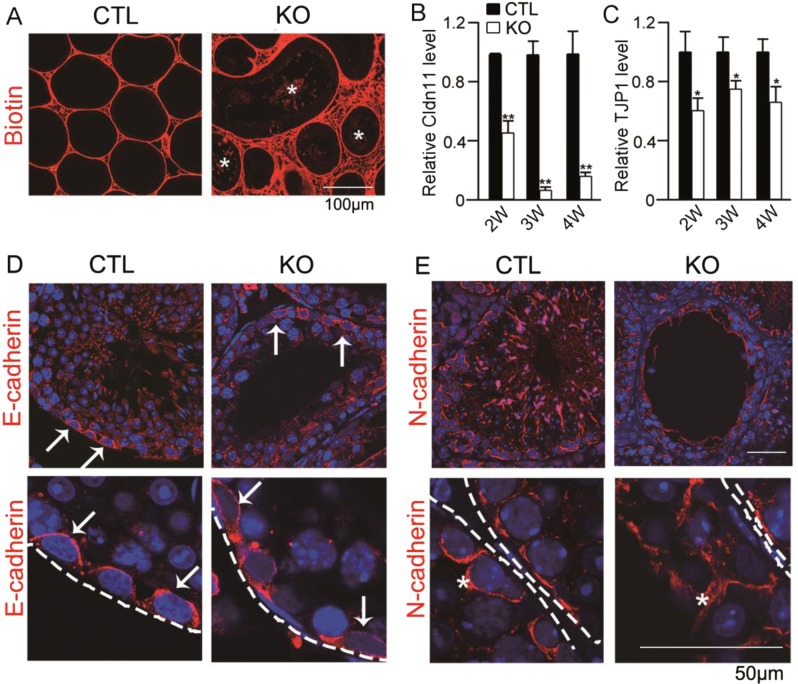
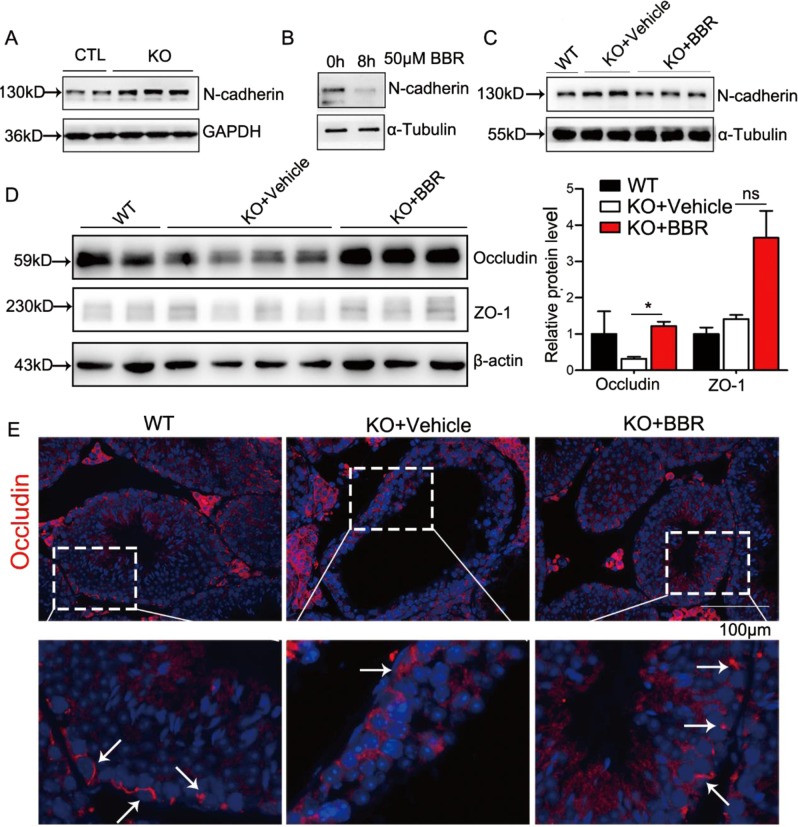
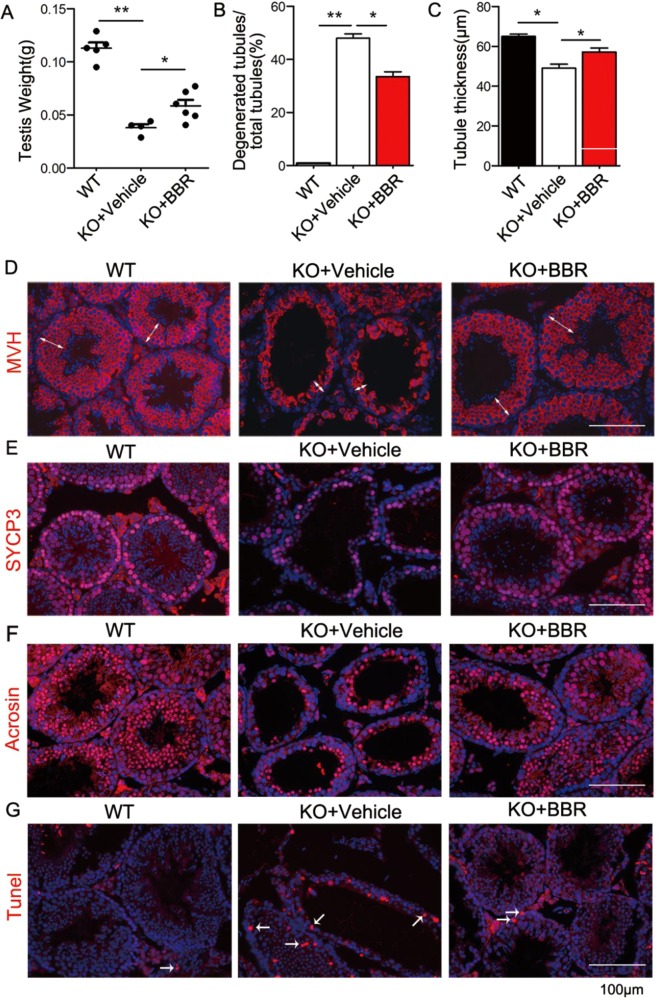
Non-obstructive azoospermia (NOA) severely affects male infertility, however, the deep mechanisms of this disease are rarely interpreted. In this study, we find that undifferentiated spermatogonial stem cells (SSCs) still exist in the basal compartment of the seminiferous tubules and the blood-testis barrier (BTB) formed by the interaction of neighbor Sertoli cells (SCs) is incomplete in NOA patients with spermatogenic maturation arrest. The adhesions between SCs and germ cells (GCs) are also broken in NOA patients. Meanwhile, the expression level of geranylgeranyl diphosphate synthase (Ggpps), a key enzyme in mevalonate metabolic pathway, is lower in NOA patients than that in obstructive azoospermia (OA) patients. After Ggpps deletion specifically in SCs, the mice are infertile and the phenotype of the SC-Ggpps-/- mice is similar to the NOA patients, where the BTB and the SC-GC adhesions are severely destroyed. Although SSCs are still found in the basal compartment of the seminiferous tubules, fewer mature spermatocyte and spermatid are found in SC-Ggpps-/- mice. Further examination suggests that the defect is mediated by the aberrant protein isoprenylation of RhoA and Ras family after Ggpps deletion. The exciting finding is that when the knockout mice are injected with berberine, the abnormal cell adhesions are ameliorated and spermatogenesis is partially restored. Our data suggest that the reconstruction of disrupted BTB is an effective treatment strategy for NOA patients with spermatogenic maturation arrest and hypospermatogenesis.
Conflict of interest statement
The authors declare that they have no conflicts of interest.
Figures



References
-
- Enders GC, Millette CF. Pachytene spermatocyte and round spermatid binding to Sertoli cells in vitro. J. Cell Sci. 1988;90(Part 1):105–114. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
