

Striatal circuits for reward learning and decision-making
- PMID: 31171839
- PMCID: PMC7231228
- DOI: 10.1038/s41583-019-0189-2
Striatal circuits for reward learning and decision-making
Abstract
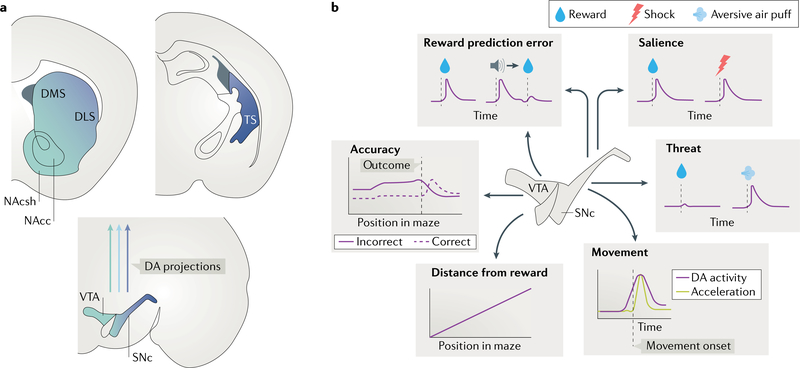
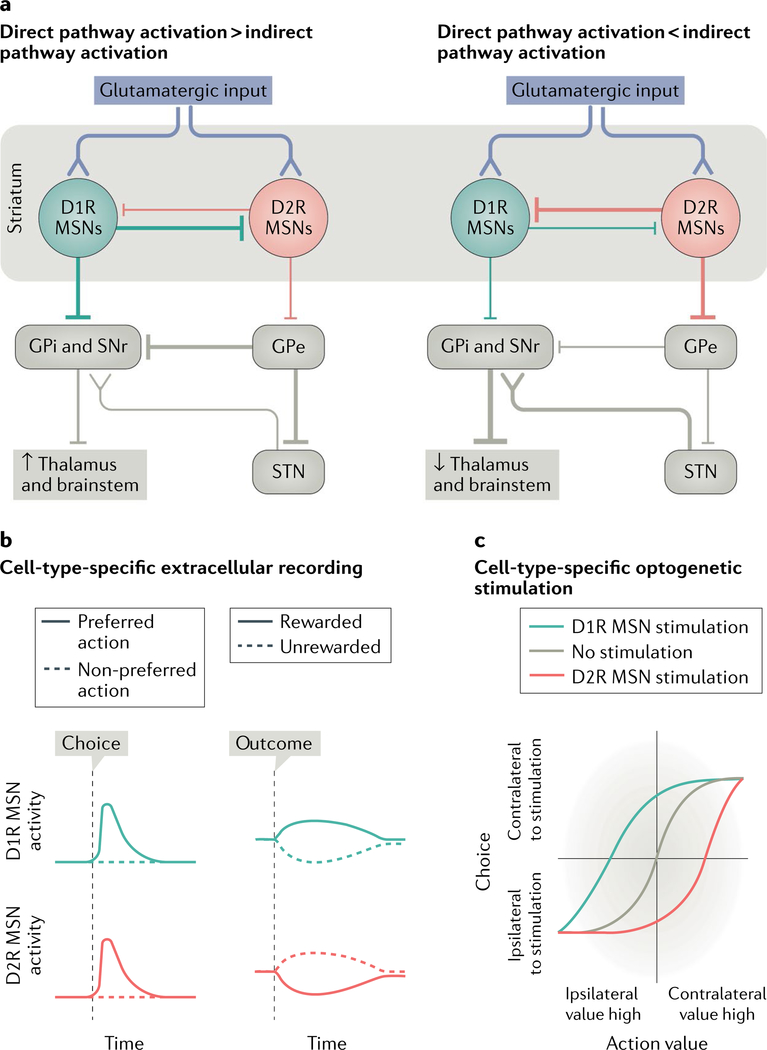
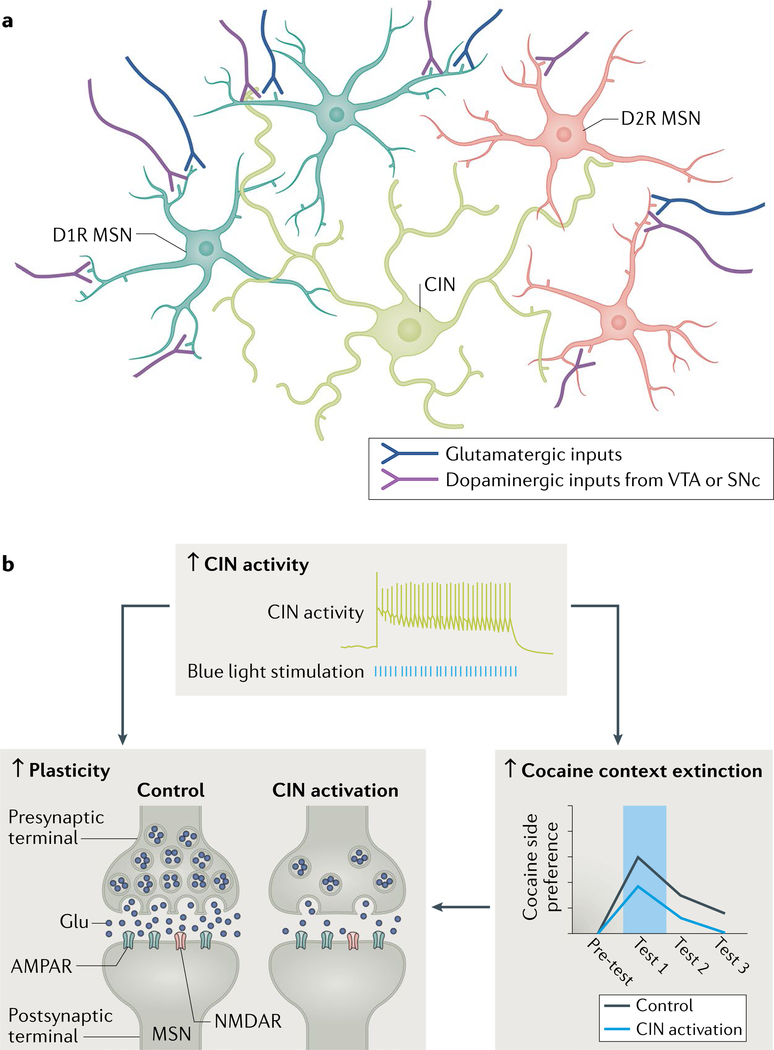
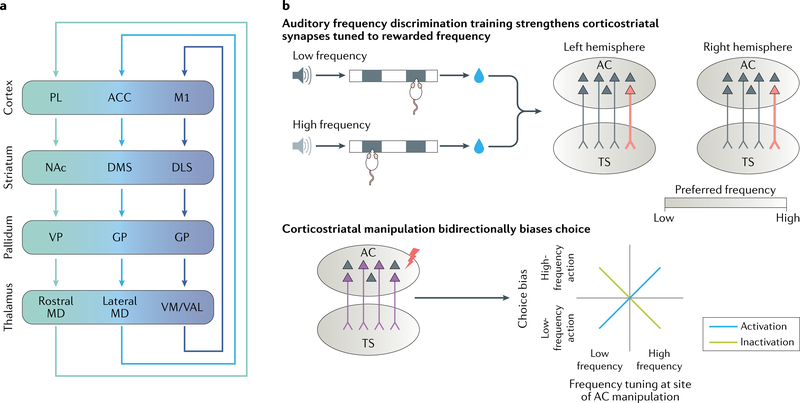
The striatum is essential for learning which actions lead to reward and for implementing those actions. Decades of experimental and theoretical work have led to several influential theories and hypotheses about how the striatal circuit mediates these functions. However, owing to technical limitations, testing these hypotheses rigorously has been difficult. In this Review, we briefly describe some of the classic ideas of striatal function. We then review recent studies in rodents that take advantage of optical and genetic methods to test these classic ideas by recording and manipulating identified cell types within the circuit. This new body of work has provided experimental support of some longstanding ideas about the striatal circuit and has uncovered critical aspects of the classic view that are incorrect or incomplete.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
-
- Alexander GE, DeLong MR & Strick PL Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu. Rev. Neurosci 9, 357–381 (1986). - PubMed
-
- Alexander GE & Crutcher MD Functional architecture of basal ganglia circuits: neural substrates of parallel processing. Trends Neurosci. 13, 266–271 (1990). - PubMed
-
- Gerfen CR & Bolam JP in Handbook of Basal Ganglia Structure and Function 2nd edn Vol. 24 (eds Steiner H & Tseng KY) 3–32 (Elsevier, 2016).
-
- Loopuijt LD & van der Kooy D Organization of the striatum: collateralization of its efferent axons. Brain Res. 348, 86–99 (1985). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
