

α-Ketoglutarate inhibits autophagy
- PMID: 31173576
- PMCID: PMC6594794
- DOI: 10.18632/aging.102001
α-Ketoglutarate inhibits autophagy
Abstract
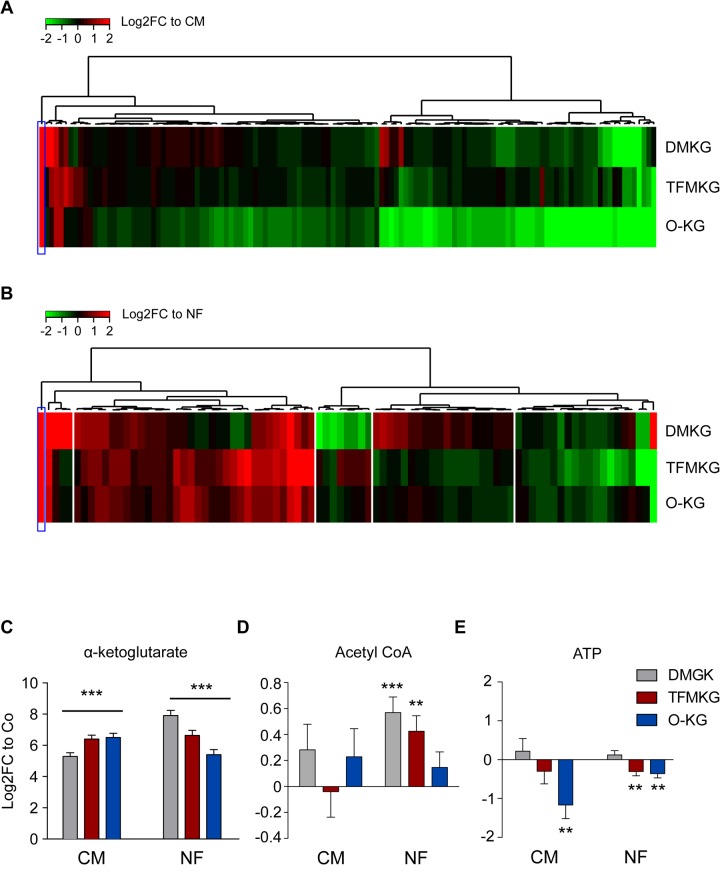
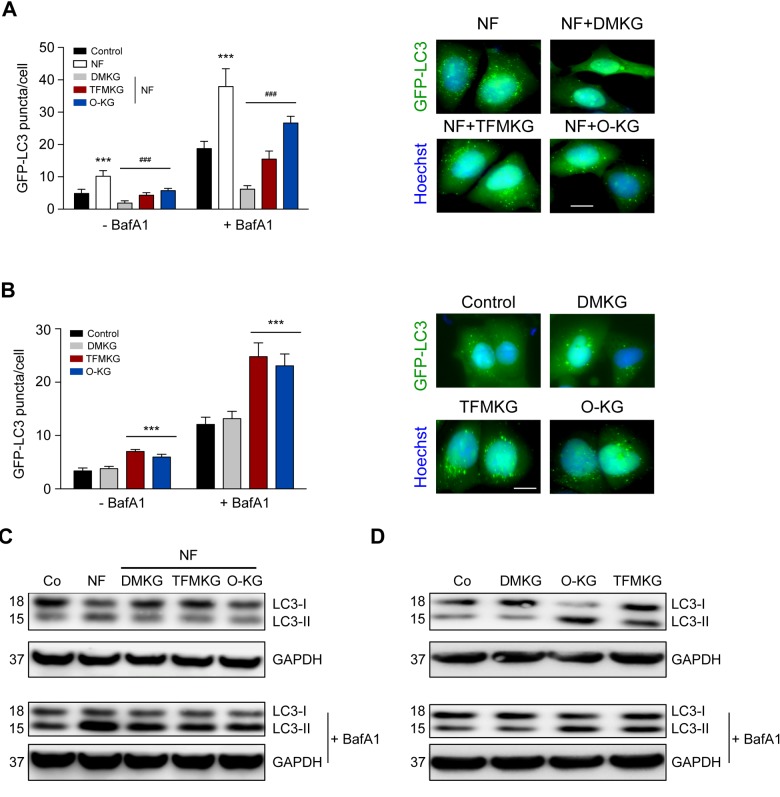
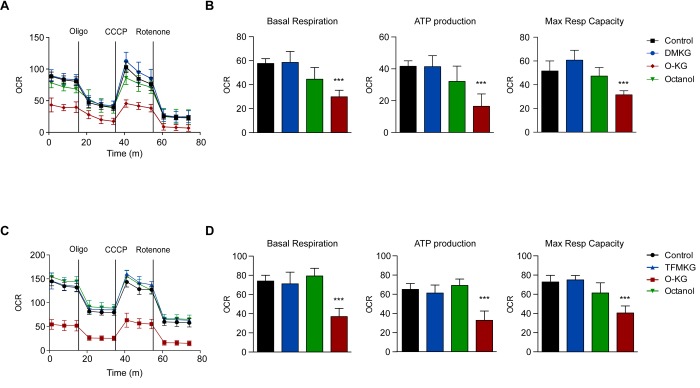
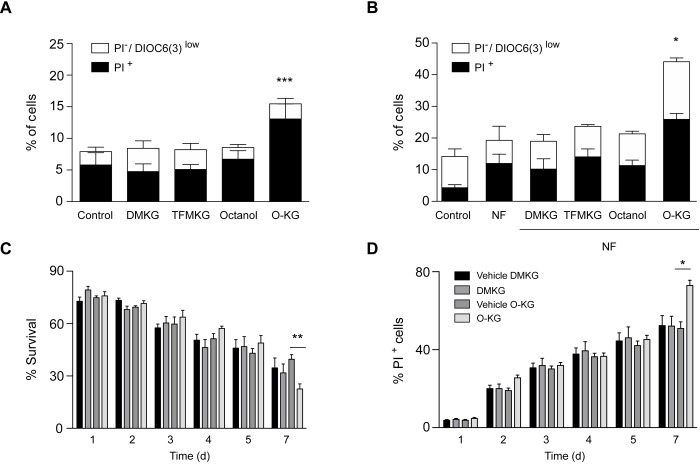
The metabolite α-ketoglutarate is membrane-impermeable, meaning that it is usually added to cells in the form of esters such as dimethyl -ketoglutarate (DMKG), trifluoromethylbenzyl α-ketoglutarate (TFMKG) and octyl α-ketoglutarate (O-KG). Once these compounds cross the plasma membrane, they are hydrolyzed by esterases to generate α-ketoglutarate, which remains trapped within cells. Here, we systematically compared DMKG, TFMKG and O-KG for their metabolic and functional effects. All three compounds similarly increased the intracellular levels of α-ketoglutarate, yet each of them had multiple effects on other metabolites that were not shared among the three agents, as determined by mass spectrometric metabolomics. While all three compounds reduced autophagy induced by culture in nutrient-free conditions, TFMKG and O-KG (but not DMKG) caused an increase in baseline autophagy in cells cultured in complete medium. O-KG (but neither DMKG nor TFMK) inhibited oxidative phosphorylation and exhibited cellular toxicity. Altogether, these results support the idea that intracellular α-ketoglutarate inhibits starvation-induced autophagy and that it has no direct respiration-inhibitory effect.
Keywords: Krebs cycle; acetyl CoA; aging; cell death; metabolomics; mitochondria.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
