

Upregulation of MIF as a defense mechanism and a biomarker of Alzheimer's disease
- PMID: 31174614
- PMCID: PMC6555932
- DOI: 10.1186/s13195-019-0508-x
Upregulation of MIF as a defense mechanism and a biomarker of Alzheimer's disease
Abstract
Background: Macrophage migration inhibitory factor (MIF) is a pro-inflammatory cytokine. Chronic inflammation induced by amyloid β proteins (Aβ) is one prominent neuropathological feature in Alzheimer's disease (AD) brain.
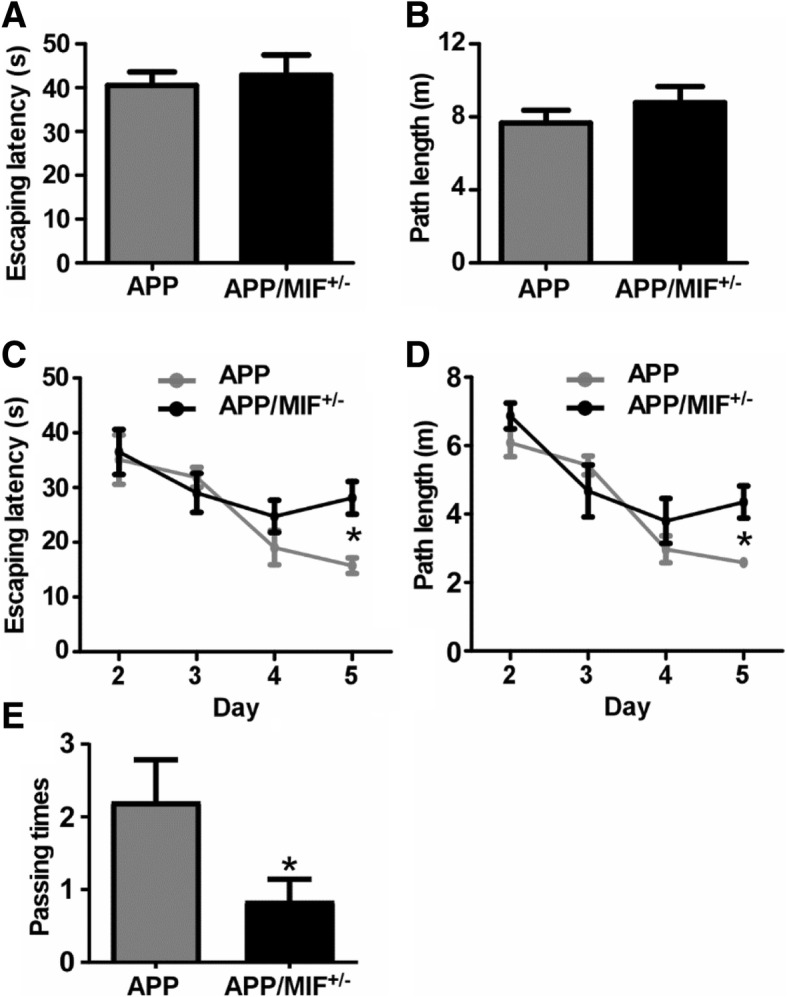
Methods: Elisa, Western blot, and immunohistochemical staining analysis were performed to examine the level of MIF protein in CSF and brain tissues. MTT and LDH assays were used to examine the neurotoxicity, and the Morris Water Maze test was performed to examine the cognitive function in the MIF+/-/APP23 transgenic mice.
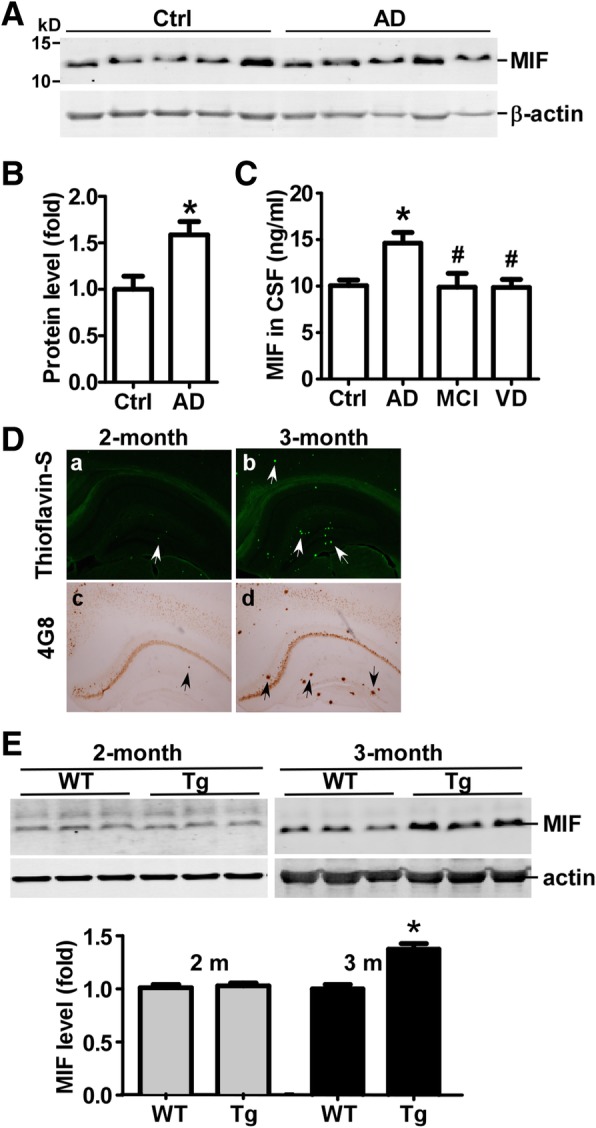
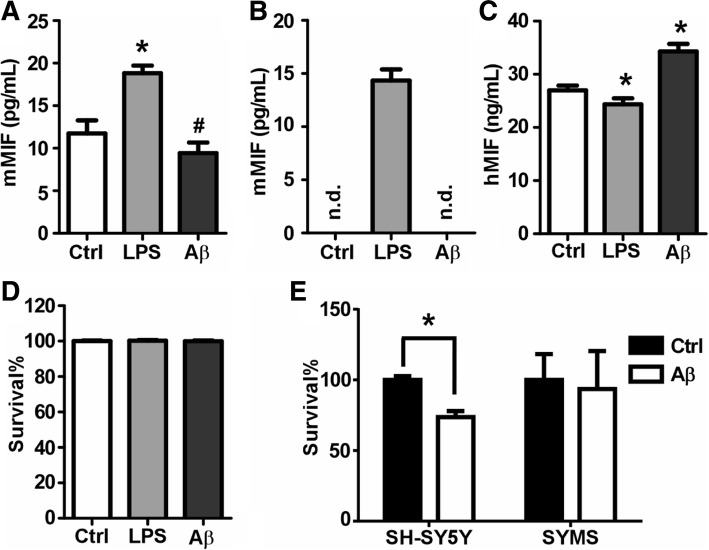
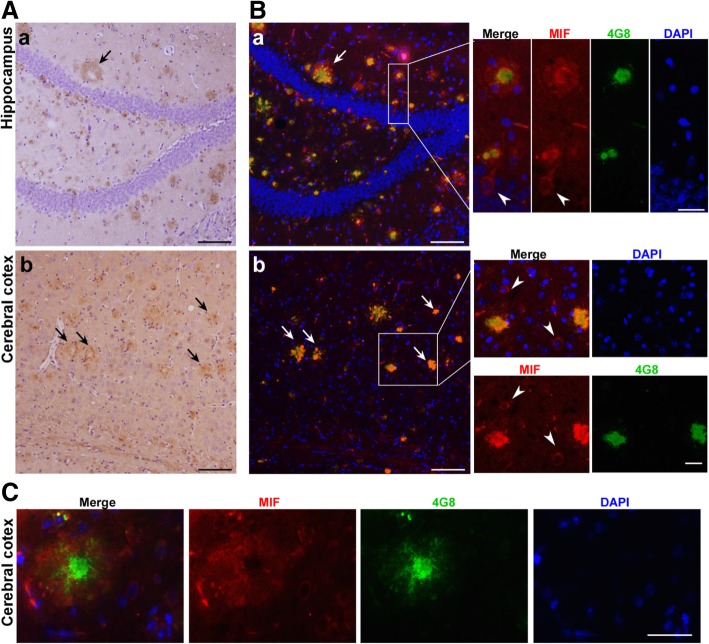
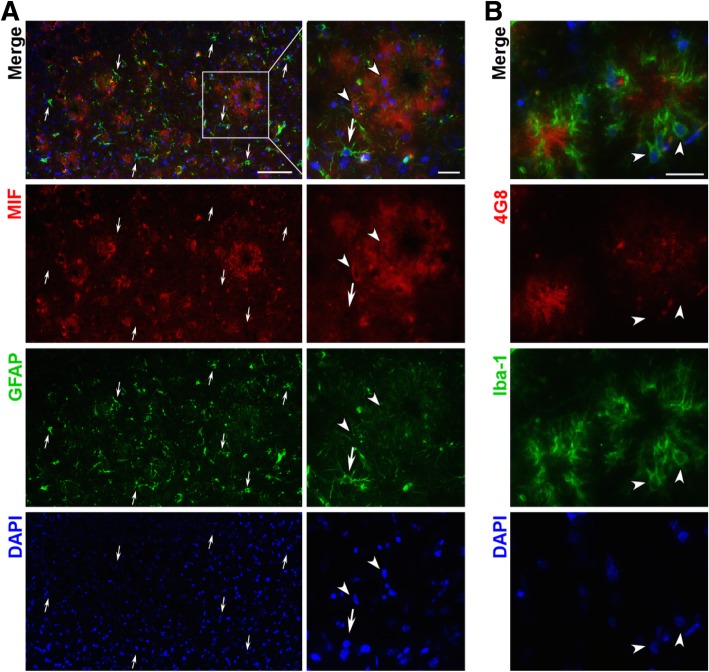
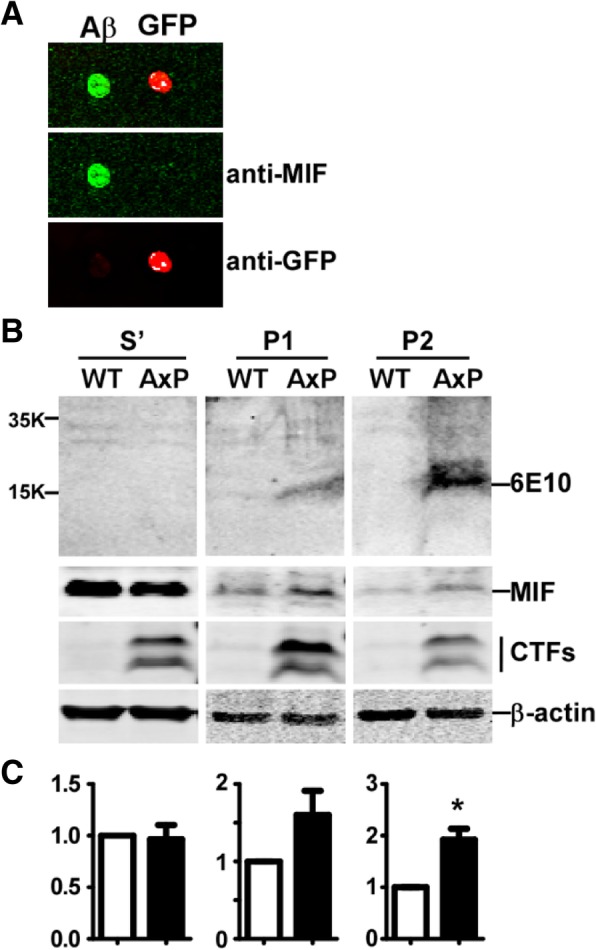
Results: MIF expression was upregulated in the brain of AD patients and AD model mice. Elevated MIF concentration was detected in the cerebrospinal fluid of AD patients but not in that of the patients suffering from mild cognitive impairment and vascular dementia. Reduced MIF expression impaired learning and memory in the AD model mice. MIF expression largely associates with Aβ deposits and microglia. The binding assay revealed a direct association between MIF and Aβ oligomers. Neurons instead of glial cells were responsible for the secretion of MIF upon stimulation by Aβ oligomers. In addition, overexpression of MIF significantly protected neuronal cells from Aβ-induced cytotoxicity.
Conclusion: Our study suggests that neuronal secretion of MIF may serve as a defense mechanism to compensate for declined cognitive function in AD, and increased MIF level could be a potential AD biomarker.
Keywords: Alzheimer’s disease; Amyloid; Cognitive impairment; MIF; Microglia; Neuronal toxicity.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
