

Regional Heterogeneity in Gene Expression, Regulation, and Coherence in the Frontal Cortex and Hippocampus across Development and Schizophrenia
- PMID: 31174959
- PMCID: PMC7000204
- DOI: 10.1016/j.neuron.2019.05.013
Regional Heterogeneity in Gene Expression, Regulation, and Coherence in the Frontal Cortex and Hippocampus across Development and Schizophrenia
Abstract
The hippocampus formation, although prominently implicated in schizophrenia pathogenesis, has been overlooked in large-scale genomics efforts in the schizophrenic brain. We performed RNA-seq in hippocampi and dorsolateral prefrontal cortices (DLPFCs) from 551 individuals (286 with schizophrenia). We identified substantial regional differences in gene expression and found widespread developmental differences that were independent of cellular composition. We identified 48 and 245 differentially expressed genes (DEGs) associated with schizophrenia within the hippocampus and DLPFC, with little overlap between the brain regions. 124 of 163 (76.6%) of schizophrenia GWAS risk loci contained eQTLs in any region. Transcriptome-wide association studies in each region identified many novel schizophrenia risk features that were brain region-specific. Last, we identified potential molecular correlates of in vivo evidence of altered prefrontal-hippocampal functional coherence in schizophrenia. These results underscore the complexity and regional heterogeneity of the transcriptional correlates of schizophrenia and offer new insights into potentially causative biology.
Keywords: RNA-seq; TWAS; brain; development; eQTL; human; regional coherence; schizophrenia.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The following BrainSeq Consortium members have competing interests. M.M., T.S., K.T., and D.J.H. are employees of Astellas Pharma. N.J.B. and A.J.C. are employees of AstraZeneca. D.A.C., J.N.C., C.L.A.R., B.J.E., P.J.E., D.C.A., Y. Li, Y. Liu, K.M., B.B.M., J.E.S., and H.W. are employees of Eli Lilly and Company. M.F., D.H., and H.K. are employees of Janssen Research & Development LLC and Johnson and Johnson. M.D. and L.F. are employees of H. Lundbeck A/S. T.K.-T. and D.M. are employees of F. Hoffmann-La Roche. P.O., S.X., and J.Q. are former employees of Pfizer.
Figures
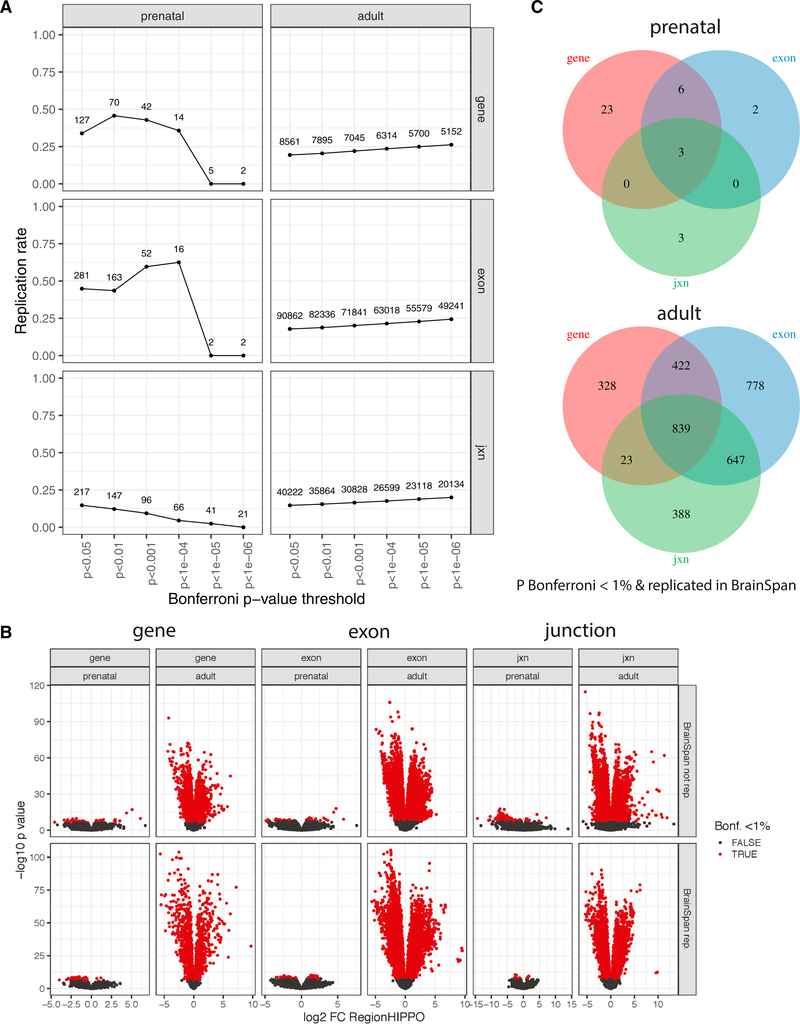
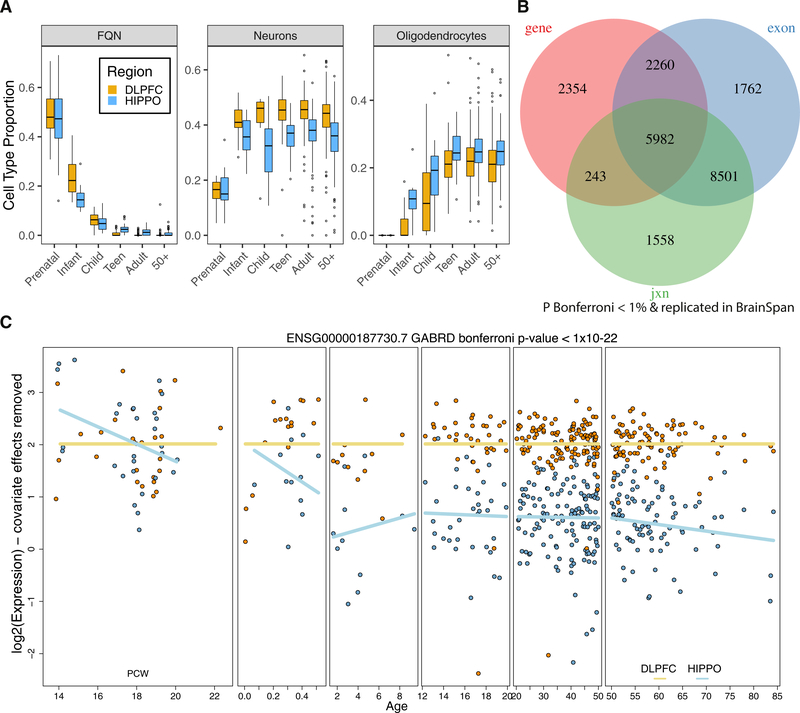
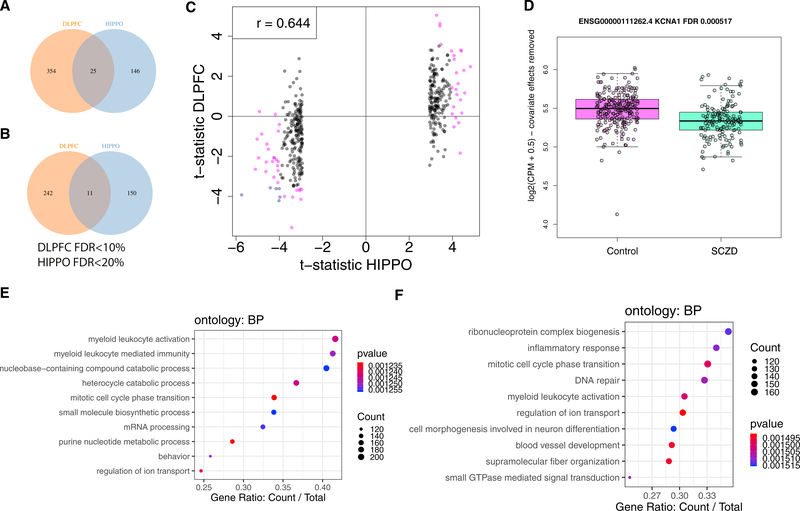
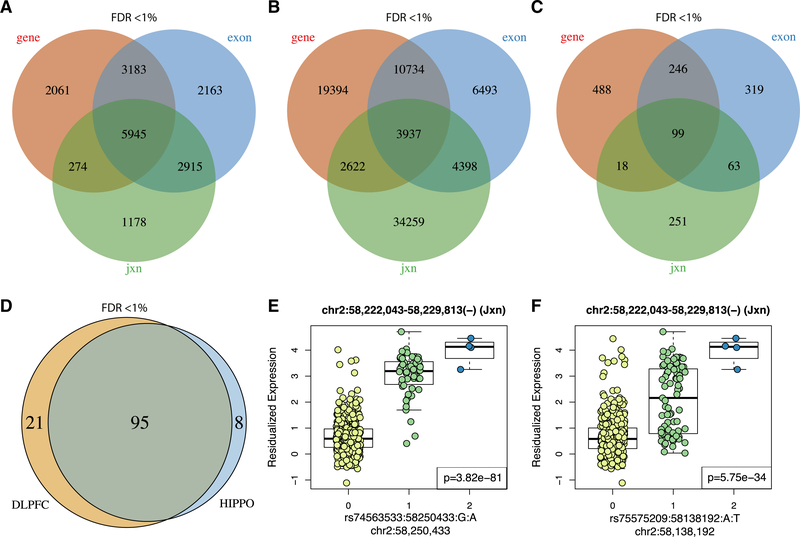
References
-
- Babraham Bioinformatics (2016). FastQC (Babraham Institute; ). https://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
