

Kinetics of Xist-induced gene silencing can be predicted from combinations of epigenetic and genomic features
- PMID: 31175153
- PMCID: PMC6633258
- DOI: 10.1101/gr.245027.118
Kinetics of Xist-induced gene silencing can be predicted from combinations of epigenetic and genomic features
Abstract
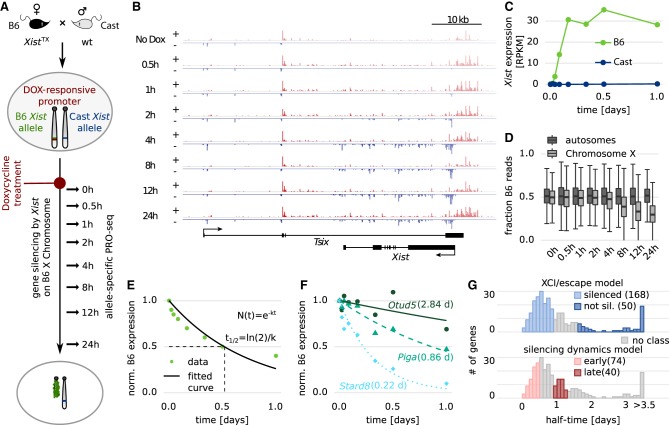
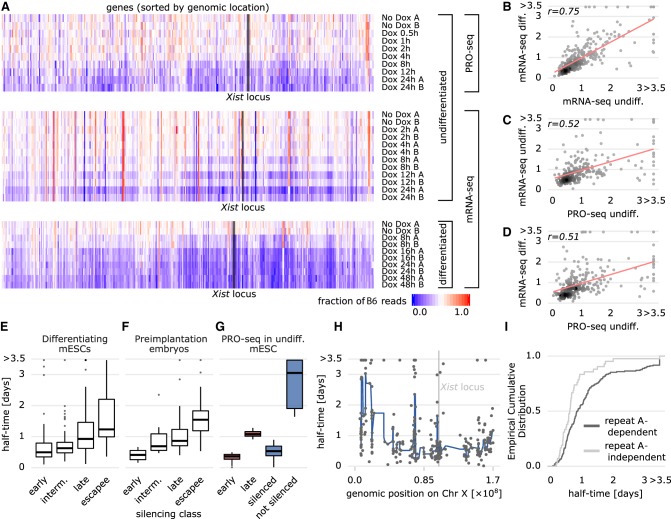
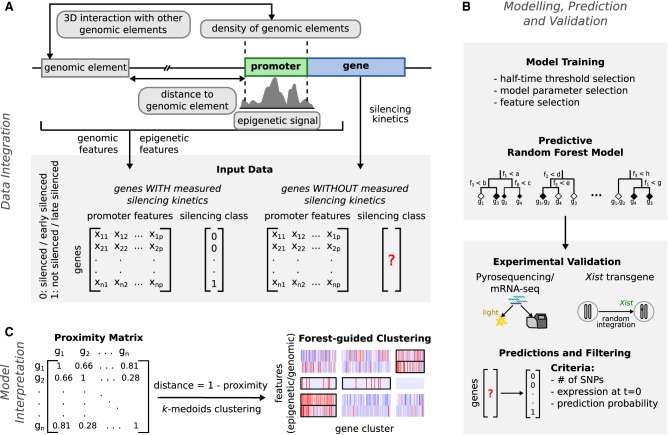
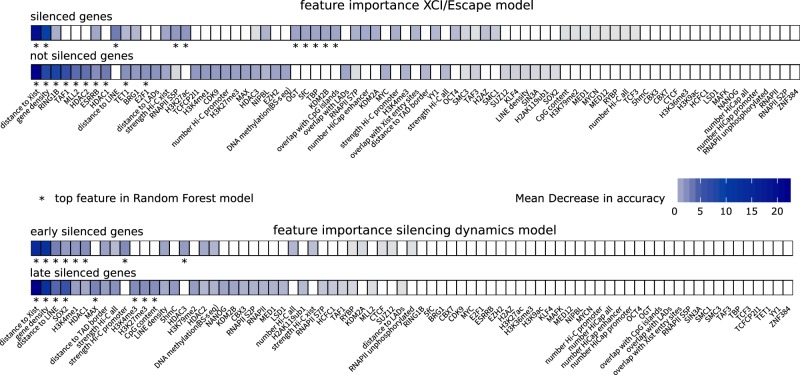
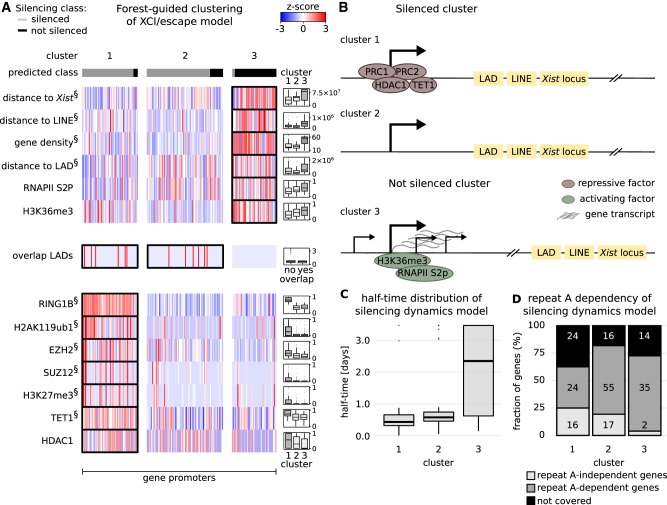
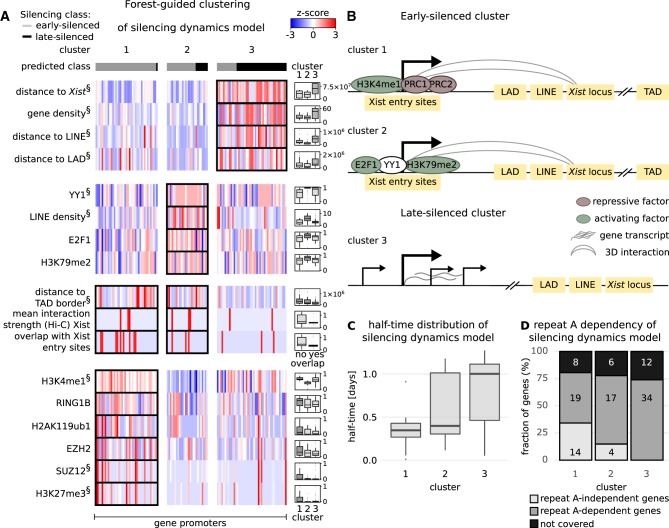
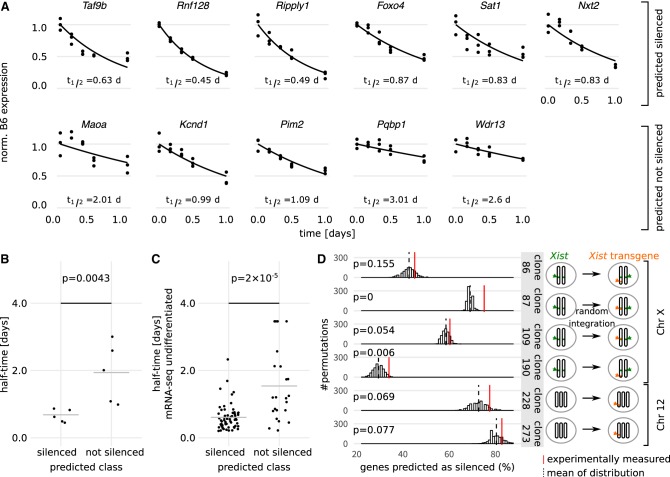
To initiate X-Chromosome inactivation (XCI), the long noncoding RNA Xist mediates chromosome-wide gene silencing of one X Chromosome in female mammals to equalize gene dosage between the sexes. The efficiency of gene silencing is highly variable across genes, with some genes even escaping XCI in somatic cells. A gene's susceptibility to Xist-mediated silencing appears to be determined by a complex interplay of epigenetic and genomic features; however, the underlying rules remain poorly understood. We have quantified chromosome-wide gene silencing kinetics at the level of the nascent transcriptome using allele-specific Precision nuclear Run-On sequencing (PRO-seq). We have developed a Random Forest machine-learning model that can predict the measured silencing dynamics based on a large set of epigenetic and genomic features and tested its predictive power experimentally. The genomic distance to the Xist locus, followed by gene density and distance to LINE elements, are the prime determinants of the speed of gene silencing. Moreover, we find two distinct gene classes associated with different silencing pathways: a class that requires Xist-repeat A for silencing, which is known to activate the SPEN pathway, and a second class in which genes are premarked by Polycomb complexes and tend to rely on the B repeat in Xist for silencing, known to recruit Polycomb complexes during XCI. Moreover, a series of features associated with active transcriptional elongation and chromatin 3D structure are enriched at rapidly silenced genes. Our machine-learning approach can thus uncover the complex combinatorial rules underlying gene silencing during X inactivation.
© 2019 Barros de Andrade e Sousa et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases