

Spliced X-box Binding Protein 1 Stimulates Adaptive Growth Through Activation of mTOR
- PMID: 31177839
- PMCID: PMC6690809
- DOI: 10.1161/CIRCULATIONAHA.118.038924
Spliced X-box Binding Protein 1 Stimulates Adaptive Growth Through Activation of mTOR
Abstract
Background: The unfolded protein response plays versatile roles in physiology and pathophysiology. Its connection to cell growth, however, remains elusive. Here, we sought to define the role of unfolded protein response in the regulation of cardiomyocyte growth in the heart.
Methods: We used both gain- and loss-of-function approaches to genetically manipulate XBP1s (spliced X-box binding protein 1), the most conserved signaling branch of the unfolded protein response, in the heart. In addition, primary cardiomyocyte culture was used to address the role of XBP1s in cell growth in a cell-autonomous manner.
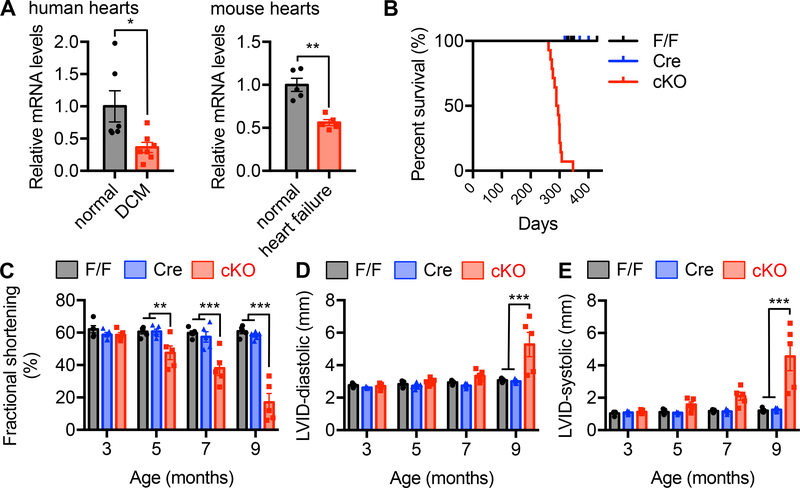
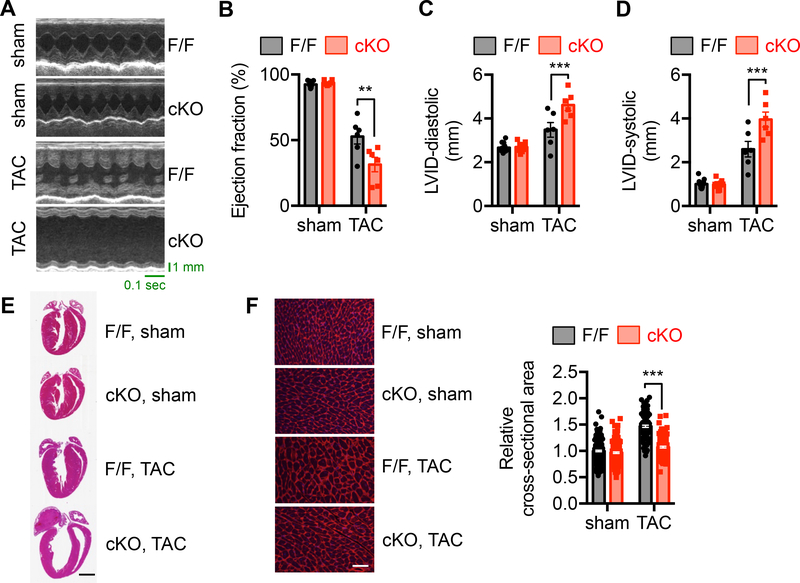
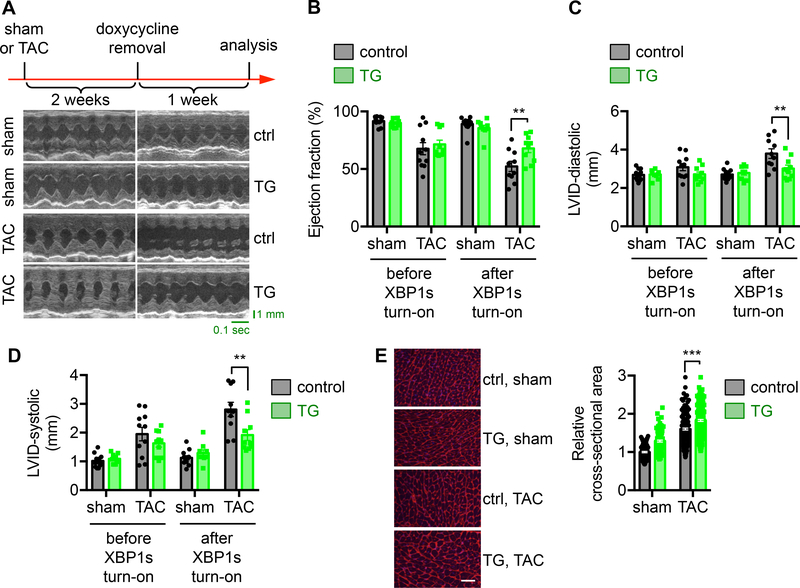
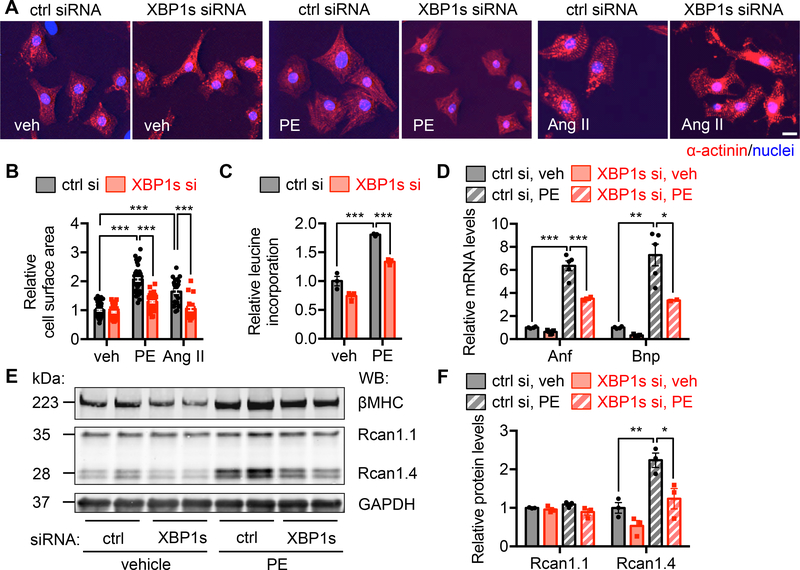
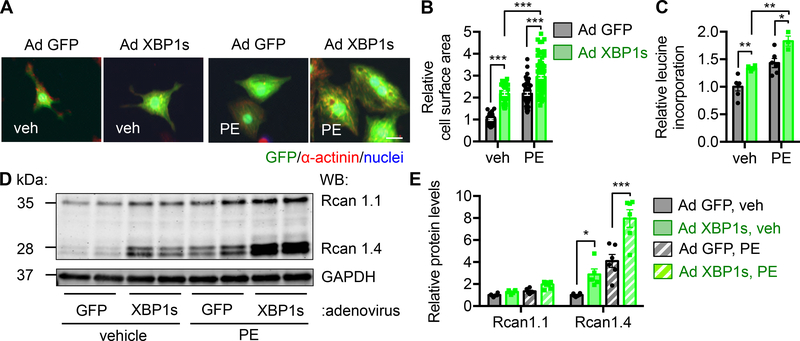
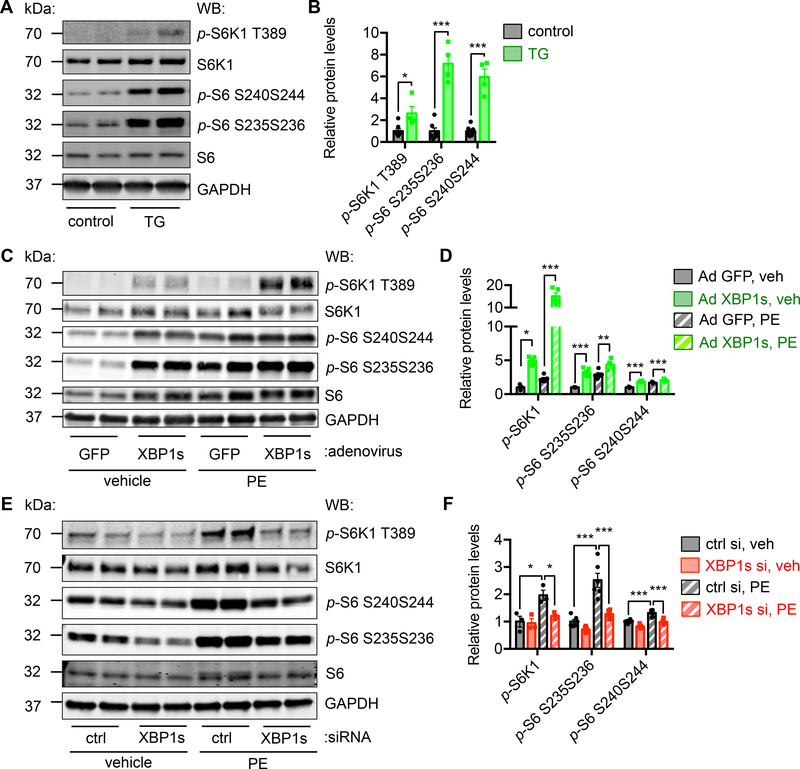
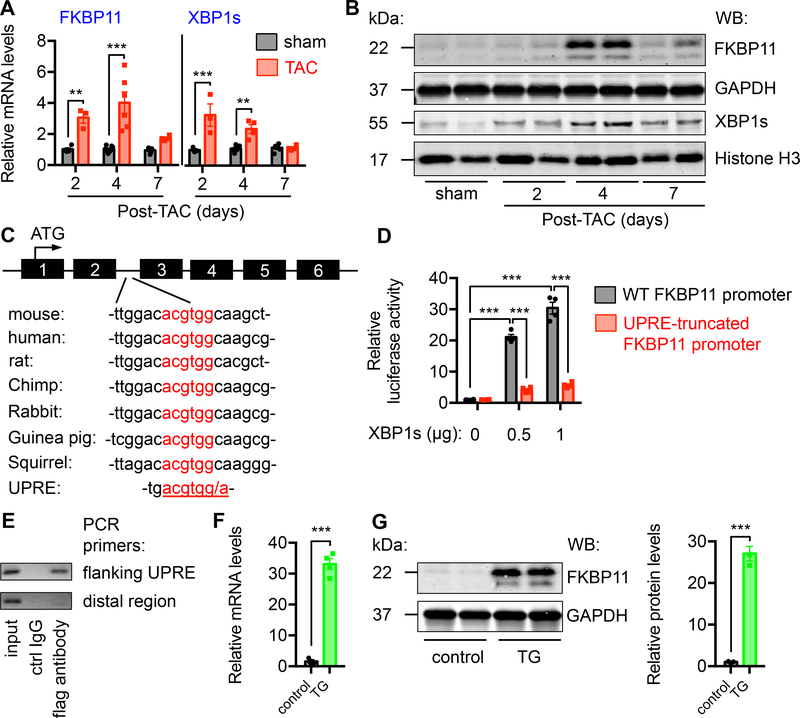
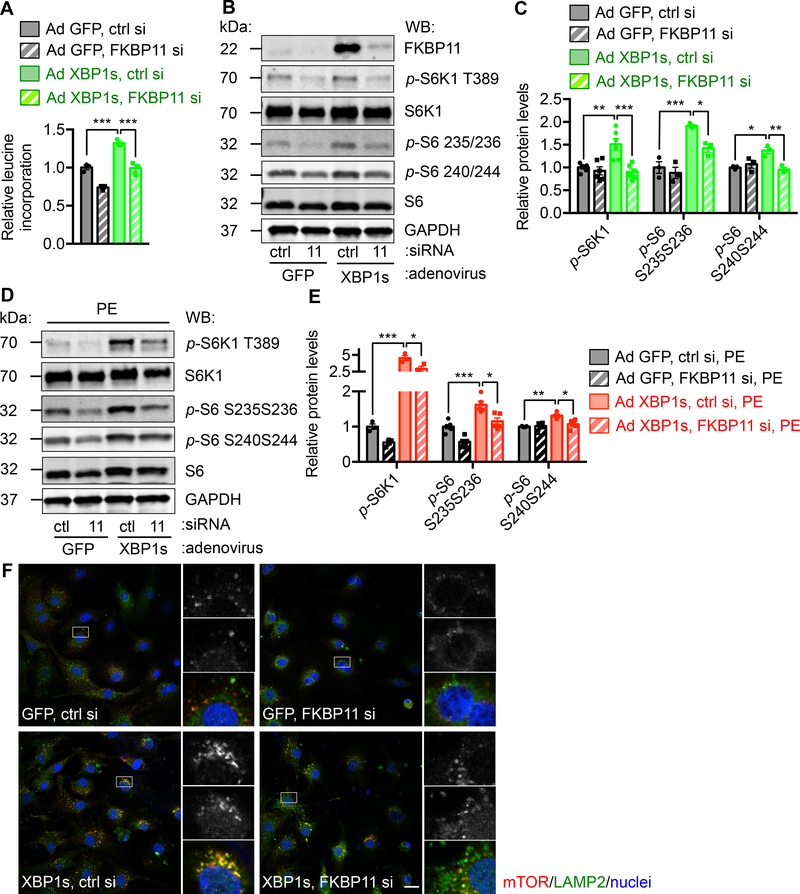
Results: We found that XBP1s expression is reduced in both human and rodent cardiac tissues under heart failure. Furthermore, deficiency of XBP1s leads to decompensation and exacerbation of heart failure progression under pressure overload. On the other hand, cardiac-restricted overexpression of XBP1s prevents the development of cardiac dysfunction. Mechanistically, we found that XBP1s stimulates adaptive cardiac growth through activation of the mechanistic target of rapamycin signaling, which is mediated via FKBP11 (FK506-binding protein 11), a novel transcriptional target of XBP1s. Moreover, silencing of FKBP11 significantly diminishes XBP1s-induced mechanistic target of rapamycin activation and adaptive cell growth.
Conclusions: Our results reveal a critical role of the XBP1s-FKBP11-mechanistic target of rapamycin axis in coupling of the unfolded protein response and cardiac cell growth regulation.
Keywords: FKBP11 protein, human; FKBP11 protein, mouse; X-box binding protein 1; heart failure; hypertrophy; mTOR protein, mouse; unfolded protein response.
Figures
References
-
- Benjamin EJ, Virani SS, Callaway CW, Chamberlain AM, Chang AR, Cheng S, Chiuve SE, Cushman M, Delling FN, Deo R, de Ferranti SD, Ferguson JF, Fornage M, Gillespie C, Isasi CR, Jimenez MC, Jordan LC, Judd SE, Lackland D, Lichtman JH, Lisabeth L, Liu S, Longenecker CT, Lutsey PL, Mackey JS, Matchar DB, Matsushita K, Mussolino ME, Nasir K, O’Flaherty M, Palaniappan LP, Pandey A, Pandey DK, Reeves MJ, Ritchey MD, Rodriguez CJ, Roth GA, Rosamond WD, Sampson UKA, Satou GM, Shah SH, Spartano NL, Tirschwell DL, Tsao CW, Voeks JH, Willey JZ, Wilkins JT, Wu JH, Alger HM, Wong SS and Muntner P. Heart Disease and Stroke Statistics-2018 Update: A Report From the American Heart Association. Circulation. 2018;137:e67–e492. - PubMed
-
- Drazner MH. The progression of hypertensive heart disease. Circulation. 2011;123:327–334. - PubMed
-
- Uhlen M, Fagerberg L, Hallstrom BM, Lindskog C, Oksvold P, Mardinoglu A, Sivertsson A, Kampf C, Sjostedt E, Asplund A, Olsson I, Edlund K, Lundberg E, Navani S, Szigyarto CA, Odeberg J, Djureinovic D, Takanen JO, Hober S, Alm T, Edqvist PH, Berling H, Tegel H, Mulder J, Rockberg J, Nilsson P, Schwenk JM, Hamsten M, von Feilitzen K, Forsberg M, Persson L, Johansson F, Zwahlen M, von Heijne G, Nielsen J and Ponten F. Proteomics. Tissue-based map of the human proteome. Science. 2015;347:1260419. - PubMed
-
- Walter P and Ron D. The unfolded protein response: from stress pathway to homeostatic regulation. Science. 2011;334:1081–1086. - PubMed
-
- Hetz C and Mollereau B. Disturbance of endoplasmic reticulum proteostasis in neurodegenerative diseases. Nat Rev Neurosci. 2014;15:233–249. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
