

Long-Term Intranasal Insulin Aspart: A Profile of Gene Expression, Memory, and Insulin Receptors in Aged F344 Rats
- PMID: 31180116
- PMCID: PMC7243593
- DOI: 10.1093/gerona/glz105
Long-Term Intranasal Insulin Aspart: A Profile of Gene Expression, Memory, and Insulin Receptors in Aged F344 Rats
Abstract
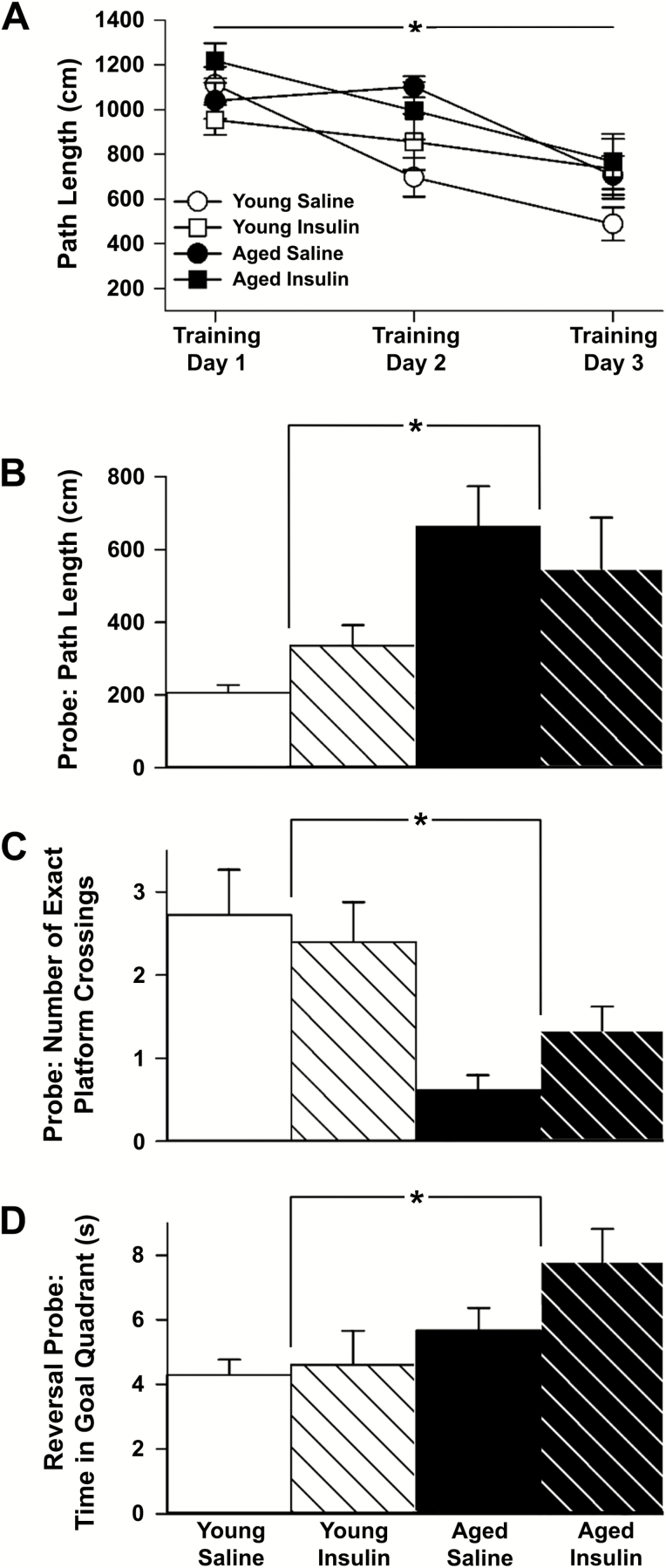
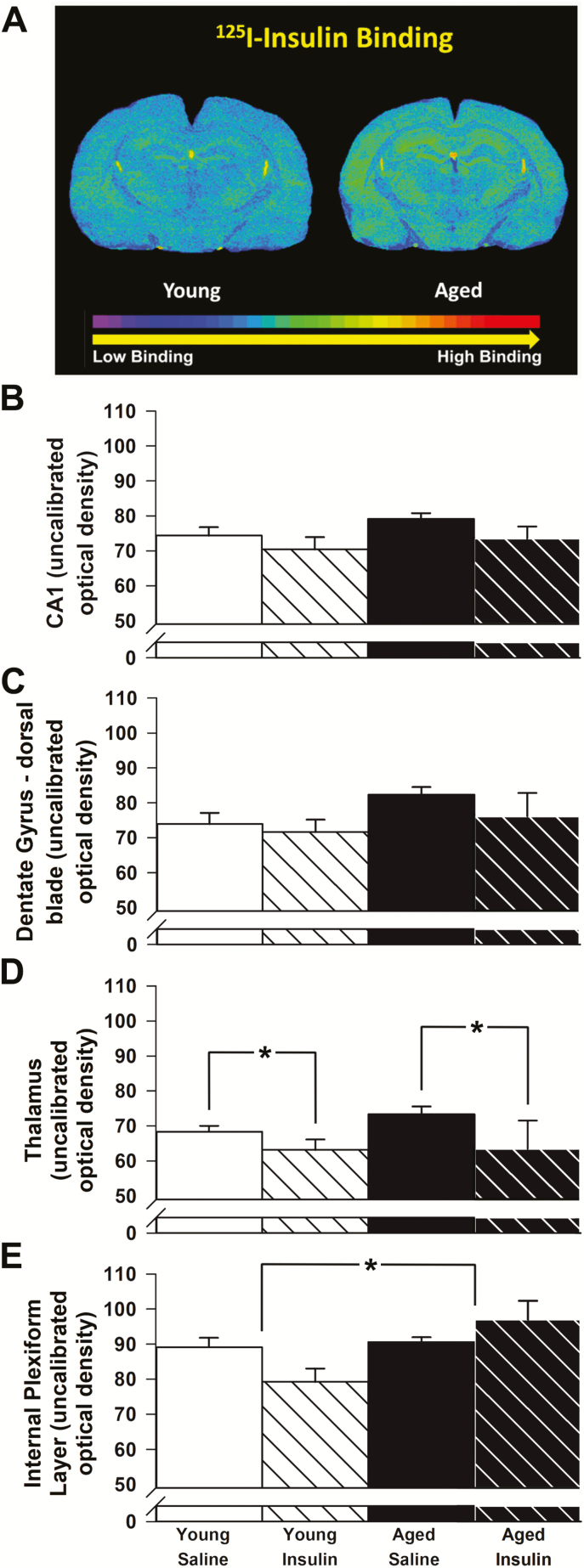
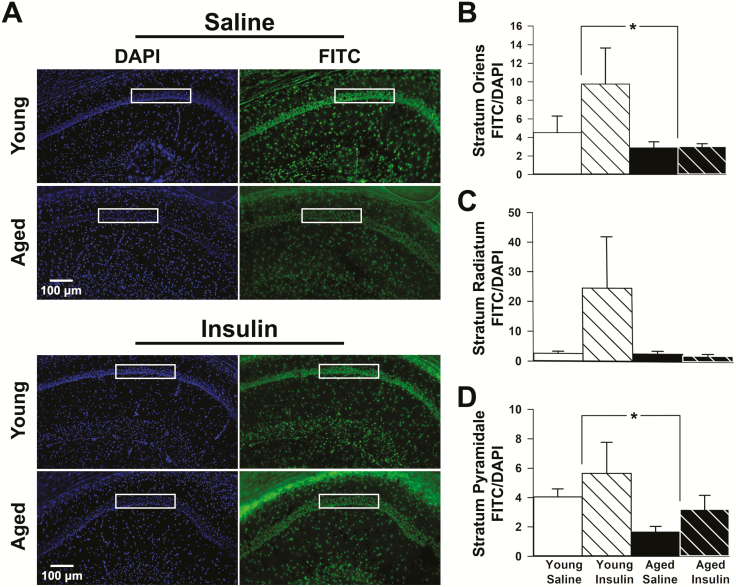
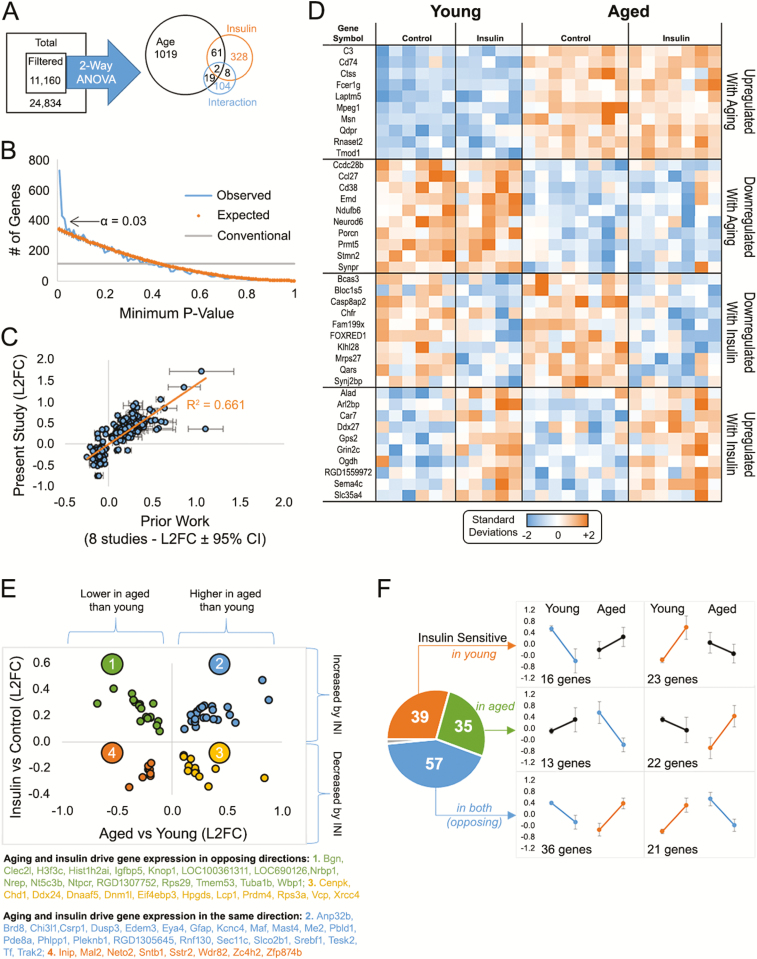
Intranasal insulin is a safe and effective method for ameliorating memory deficits associated with pathological brain aging. However, the impact of different formulations and the duration of treatment on insulin's efficacy and the cellular processes targeted by the treatment remain unclear. Here, we tested whether intranasal insulin aspart, a short-acting insulin formulation, could alleviate memory decline associated with aging and whether long-term treatment affected regulation of insulin receptors and other potential targets. Outcome variables included measures of spatial learning and memory, autoradiography and immunohistochemistry of the insulin receptor, and hippocampal microarray analyses. Aged Fischer 344 rats receiving long-term (3 months) intranasal insulin did not show significant memory enhancement on the Morris water maze task. Autoradiography results showed that long-term treatment reduced insulin binding in the thalamus but not the hippocampus. Results from hippocampal immunofluorescence revealed age-related decreases in insulin immunoreactivity that were partially offset by intranasal administration. Microarray analyses highlighted numerous insulin-sensitive genes, suggesting insulin aspart was able to enter the brain and alter hippocampal RNA expression patterns including those associated with tumor suppression. Our work provides insights into potential mechanisms of intranasal insulin and insulin resistance, and highlights the importance of treatment duration and the brain regions targeted.
Keywords: Animal model; Antiaging; Cognitive decline.
© The Author(s) 2019. Published by Oxford University Press on behalf of The Gerontological Society of America. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
References
-
- Lochhead JJ, Thorne RG. Intranasal delivery of biologics to the central nervous system. Adv Drug Deliv Rev. 2012;64:614–628. doi:10.1016/j.addr.2011.11.002 - PubMed
-
- Benedict C, Hallschmid M, Schmitz K, et al. . Intranasal insulin improves memory in humans: superiority of insulin aspart. Neuropsychopharmacology. 2007;32:239–243. doi:10.1038/sj.npp.1301193 - PubMed
-
- Born J, Lange T, Kern W, McGregor GP, Bickel U, Fehm HL. Sniffing neuropeptides: a transnasal approach to the human brain. Nat Neurosci. 2002;5:514–516. doi:10.1038/nn849 - PubMed
-
- Thorne RG, Emory CR, Ala TA, Frey WH II. Quantitative analysis of the olfactory pathway for drug delivery to the brain. Brain Res. 1995;692:278–282. doi:10.1016/0006-8993(95)00637-6 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
