

Molecular organization and dynamics of the fusion protein Gc at the hantavirus surface
- PMID: 31180319
- PMCID: PMC6609335
- DOI: 10.7554/eLife.46028
Molecular organization and dynamics of the fusion protein Gc at the hantavirus surface
Abstract
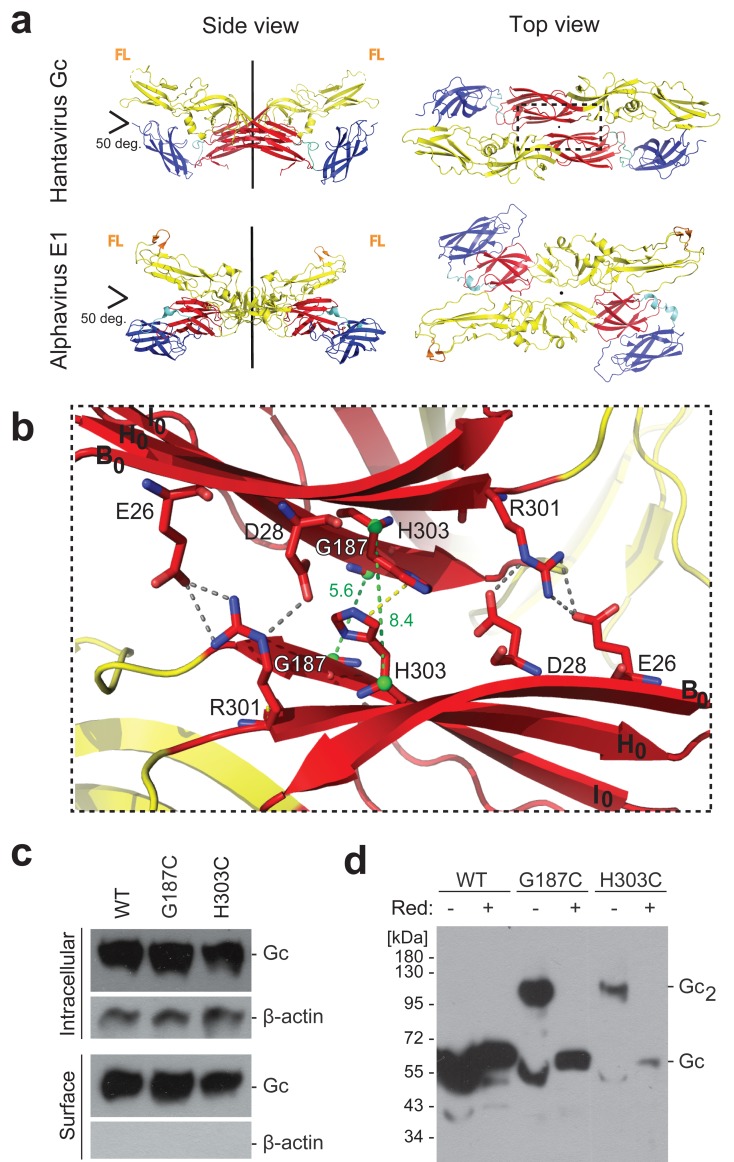
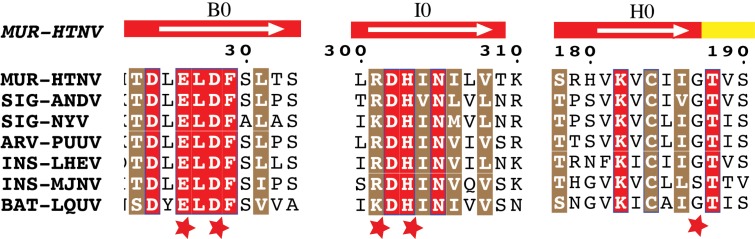
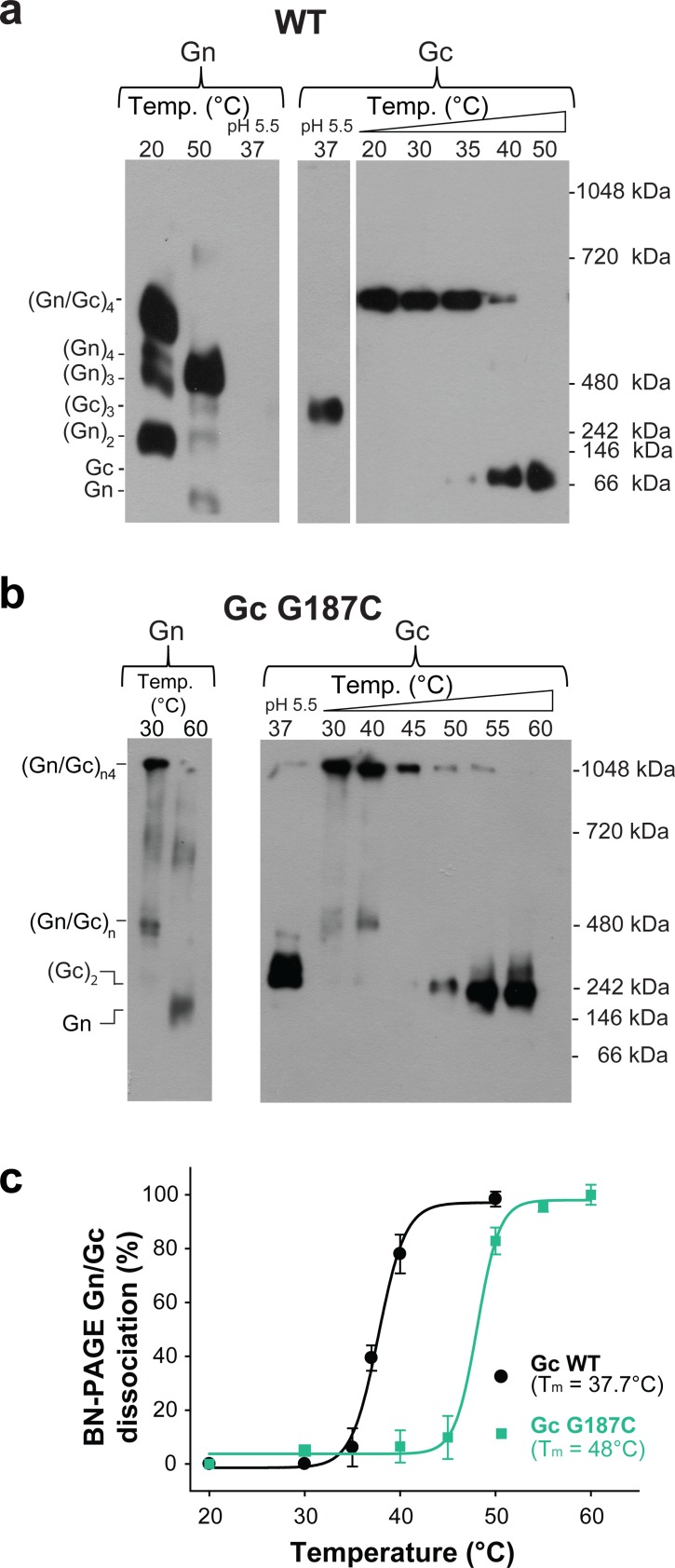
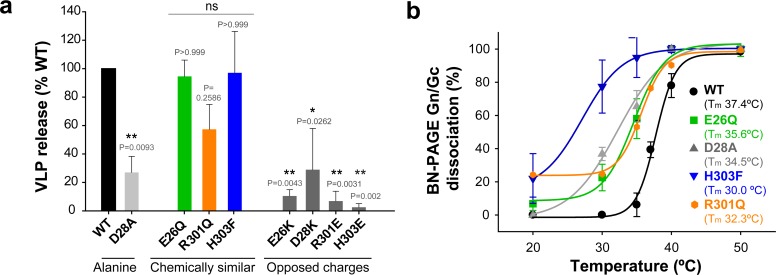
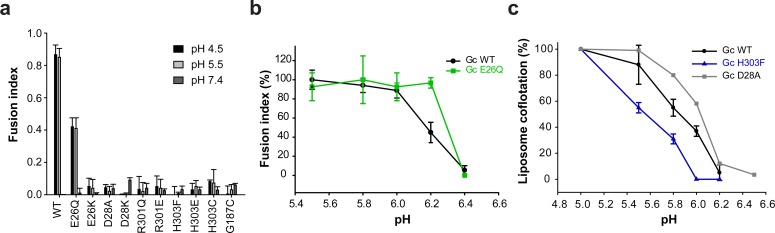
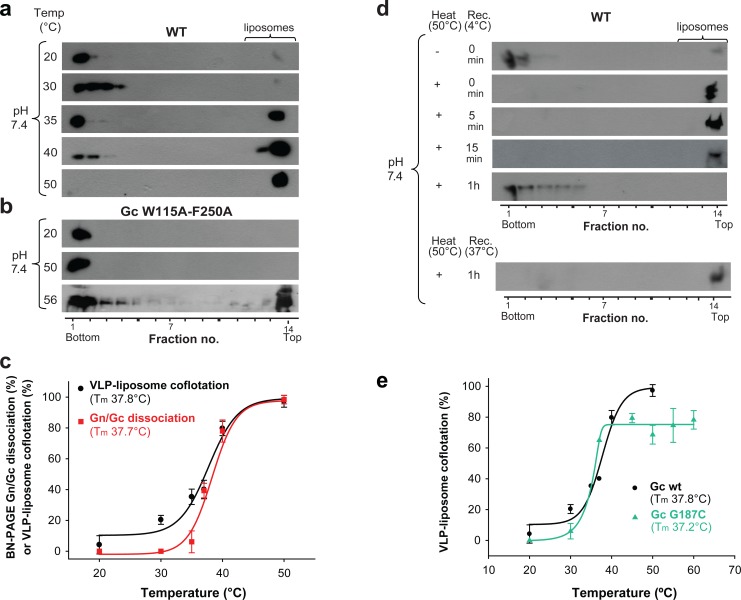
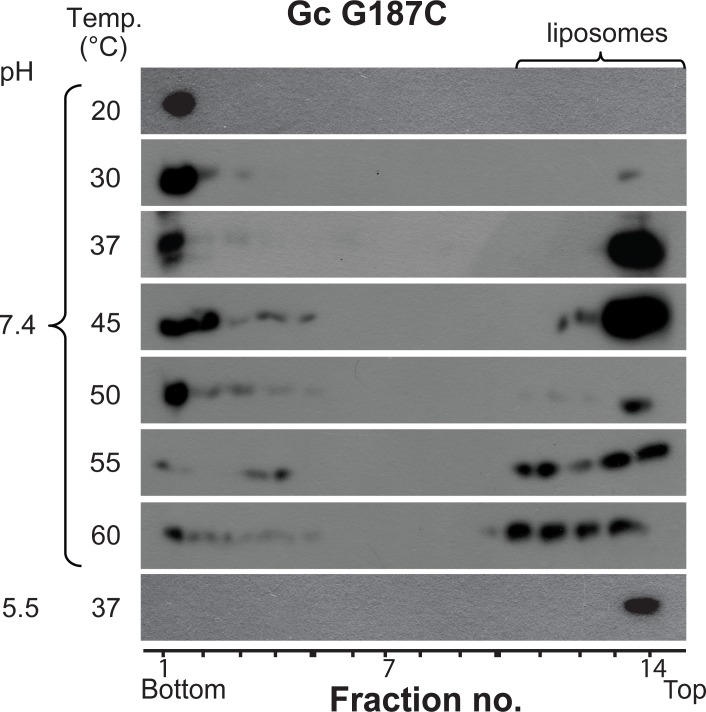
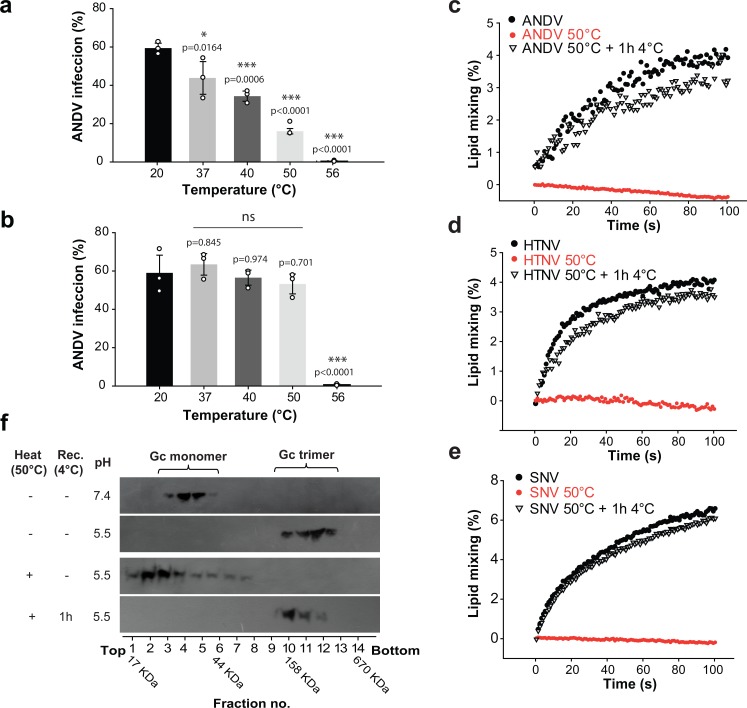
The hantavirus envelope glycoproteins Gn and Gc mediate virion assembly and cell entry, with Gc driving fusion of viral and endosomal membranes. Although the X-ray structures and overall arrangement of Gn and Gc on the hantavirus spikes are known, their detailed interactions are not. Here we show that the lateral contacts between spikes are mediated by the same 2-fold contacts observed in Gc crystals at neutral pH, allowing the engineering of disulfide bonds to cross-link spikes. Disrupting the observed dimer interface affects particle assembly and overall spike stability. We further show that the spikes display a temperature-dependent dynamic behavior at neutral pH, alternating between 'open' and 'closed' forms. We show that the open form exposes the Gc fusion loops but is off-pathway for productive Gc-induced membrane fusion and cell entry. These data also provide crucial new insights for the design of optimized Gn/Gc immunogens to elicit protective immune responses.
Keywords: Bunyavirales order; biochemistry; chemical biology; envelope proteins; infectious disease; inter-spike interface; membrane fusion; microbiology; viral breathing; virus.
© 2019, Bignon et al.
Conflict of interest statement
EB, PG, FR, NT Is named inventor on a patent application describing disulfide bonds for hantavirus spike stabilization (PCT/US19/22134). AA No competing interests declared
Figures
Similar articles
-
Hantavirus Gn and Gc envelope glycoproteins: key structural units for virus cell entry and virus assembly.Viruses. 2014 Apr 21;6(4):1801-22. doi: 10.3390/v6041801. Viruses. 2014. PMID: 24755564 Free PMC article. Review.
-
Structural Transitions of the Conserved and Metastable Hantaviral Glycoprotein Envelope.J Virol. 2017 Oct 13;91(21):e00378-17. doi: 10.1128/JVI.00378-17. Print 2017 Nov 1. J Virol. 2017. PMID: 28835498 Free PMC article.
-
Inhibition of the Hantavirus Fusion Process by Predicted Domain III and Stem Peptides from Glycoprotein Gc.PLoS Negl Trop Dis. 2016 Jul 14;10(7):e0004799. doi: 10.1371/journal.pntd.0004799. eCollection 2016 Jul. PLoS Negl Trop Dis. 2016. PMID: 27414047 Free PMC article.
-
Acidification triggers Andes hantavirus membrane fusion and rearrangement of Gc into a stable post-fusion homotrimer.J Gen Virol. 2015 Nov;96(11):3192-3197. doi: 10.1099/jgv.0.000269. Epub 2015 Aug 25. J Gen Virol. 2015. PMID: 26310672
-
The Envelope Proteins of the Bunyavirales.Adv Virus Res. 2017;98:83-118. doi: 10.1016/bs.aivir.2017.02.002. Epub 2017 Apr 8. Adv Virus Res. 2017. PMID: 28433053 Review.
Cited by
-
Broad and potently neutralizing monoclonal antibodies isolated from human survivors of New World hantavirus infection.Cell Rep. 2021 May 4;35(5):109086. doi: 10.1016/j.celrep.2021.109086. Cell Rep. 2021. PMID: 33951434 Free PMC article.
-
Human antibody recognizing a quaternary epitope in the Puumala virus glycoprotein provides broad protection against orthohantaviruses.Sci Transl Med. 2022 Mar 16;14(636):eabl5399. doi: 10.1126/scitranslmed.abl5399. Epub 2022 Mar 16. Sci Transl Med. 2022. PMID: 35294259 Free PMC article.
-
Antigenic mapping and functional characterization of human New World hantavirus neutralizing antibodies.Elife. 2023 Mar 27;12:e81743. doi: 10.7554/eLife.81743. Elife. 2023. PMID: 36971354 Free PMC article.
-
Molecular rationale for antibody-mediated targeting of the hantavirus fusion glycoprotein.Elife. 2020 Dec 22;9:e58242. doi: 10.7554/eLife.58242. Elife. 2020. PMID: 33349334 Free PMC article.
-
NMR side-chain assignments of the Crimean-Congo hemorrhagic fever virus glycoprotein n cytosolic domain.Magn Reson (Gott). 2024 Jul 8;5(2):95-101. doi: 10.5194/mr-5-95-2024. eCollection 2024. Magn Reson (Gott). 2024. PMID: 40521231 Free PMC article.
References
-
- Barriga GP, Villalón-Letelier F, Márquez CL, Bignon EA, Acuña R, Ross BH, Monasterio O, Mardones GA, Vidal SE, Tischler ND. Inhibition of the hantavirus fusion process by predicted domain III and stem peptides from glycoprotein gc. PLOS Neglected Tropical Diseases. 2016;10:e0004799. doi: 10.1371/journal.pntd.0004799. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- Fondo Nacional de Desarrollo Científico y Tecnológico FONDECYT 1181799/Comisión Nacional de Investigación Científica y Tecnológica/International
- Programa de Apoyo a Centros con Financiamiento Basal 170004 to Fundación Ciencia and Vida/Comisión Nacional de Investigación Científica y Tecnológica/International
- FONDEQUIP EQM130092 for the improvement of BSL3 of Pontificia Universidad Católica de Chile/Comisión Nacional de Investigación Científica y Tecnológica/International
- ANR-10-LABX-62-IBEID/Labex IBEID/International
- ANR-10-LABX-62-IBEID 4E AAP/Labex IBEID/International
- Infect-ERA IMI European network HantaHunt Program/Seventh Framework Programme/International
- Fondo Nacional de Desarrollo Científico y Tecnológico FONDECYT 3150695/Comisión Nacional de Investigación Científica y Tecnológica/International
- Fondo Nacional de Desarrollo Científico y Tecnológico FONDECYT 1181799/Comisión Nacional de Investigación Científica y Tecnológica/International
- Programa de Apoyo a Centros con Financiamiento Basal 170004 to Fundación Ciencia and Vida/Comisión Nacional de Investigación Científica y Tecnológica/International
- FONDEQUIP EQM130092 for the improvement of BSL3 of Pontificia Universidad Católica de Chile/Comisión Nacional de Investigación Científica y Tecnológica/International
- grant ANR-10-LABX-62-IBEID/Integrative Biology of Emerging Infectious Diseases Labex (French government´s/International
- grant ANR-10-LABX-62-IBEID 4E AAP/Labex IBEID/International
- Program/Infect-ERA IMI European network/International
- Fondo Nacional de Desarrollo Científico y Tecnológico FONDECYT 3150695/Comisión Nacional de Investigación Científica y Tecnológica/International
LinkOut - more resources
Full Text Sources
Miscellaneous
