

Sizes of actin networks sharing a common environment are determined by the relative rates of assembly
- PMID: 31181075
- PMCID: PMC6586355
- DOI: 10.1371/journal.pbio.3000317
Sizes of actin networks sharing a common environment are determined by the relative rates of assembly
Abstract
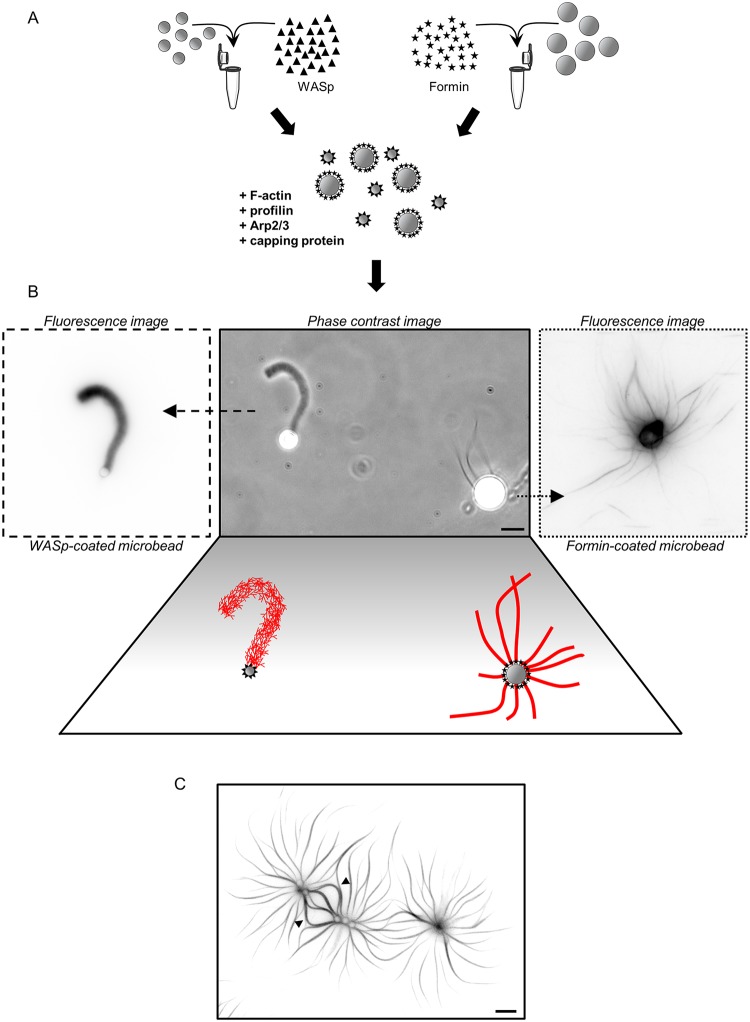
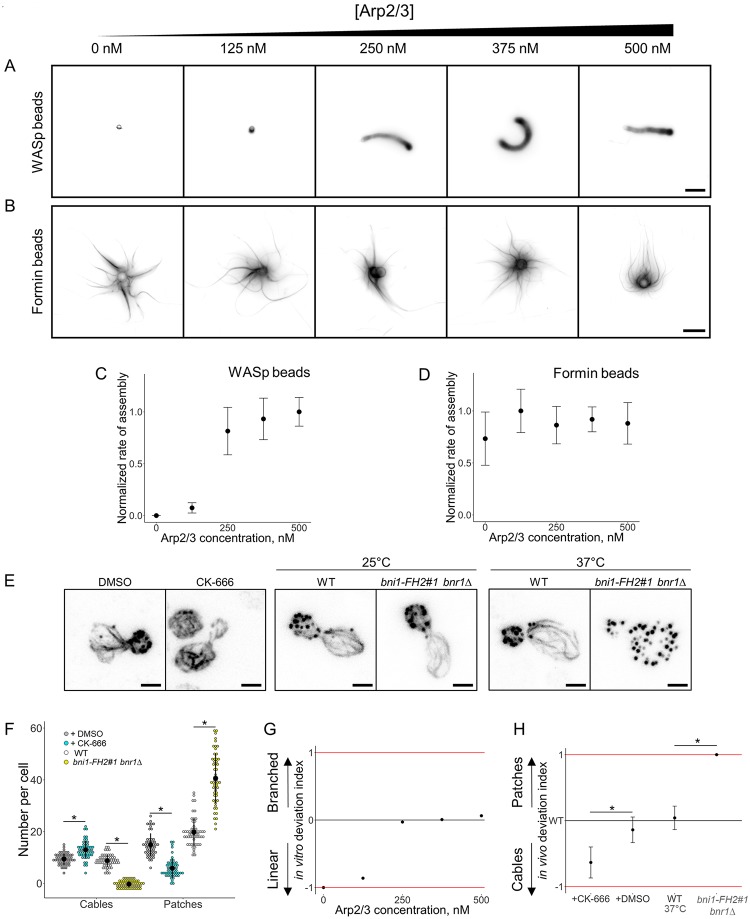
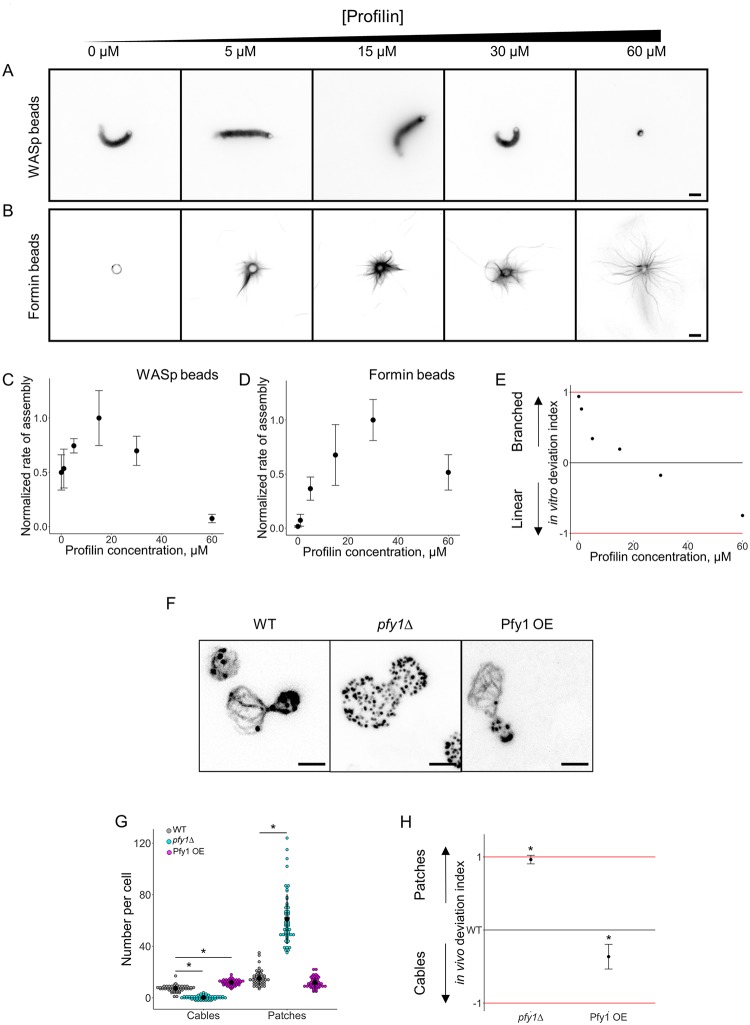
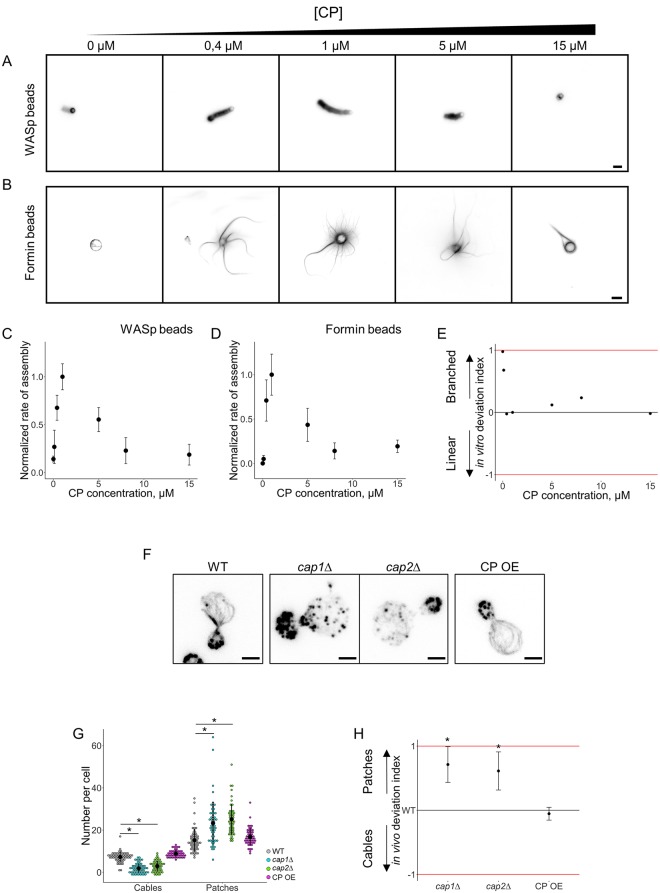
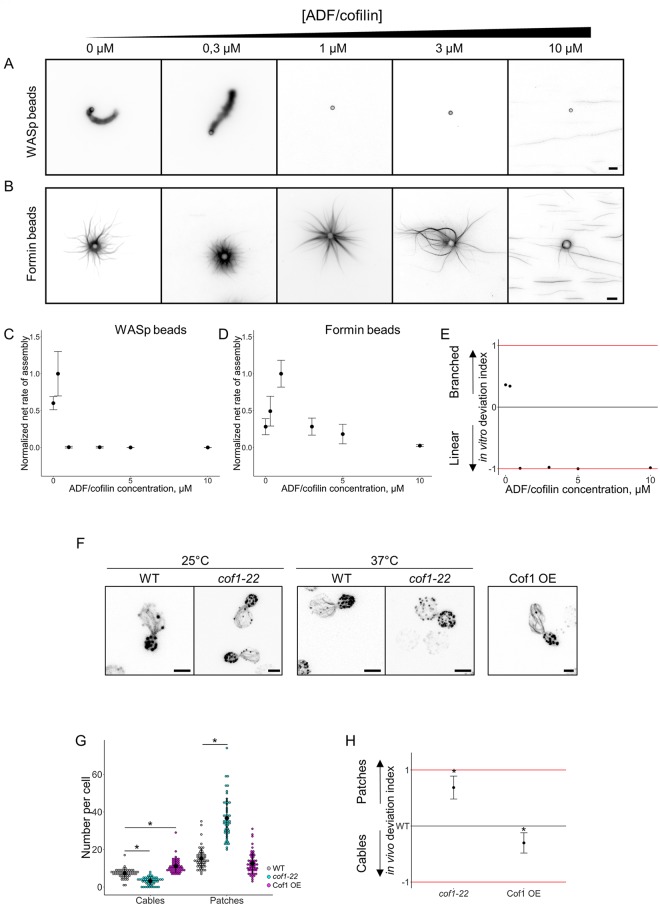
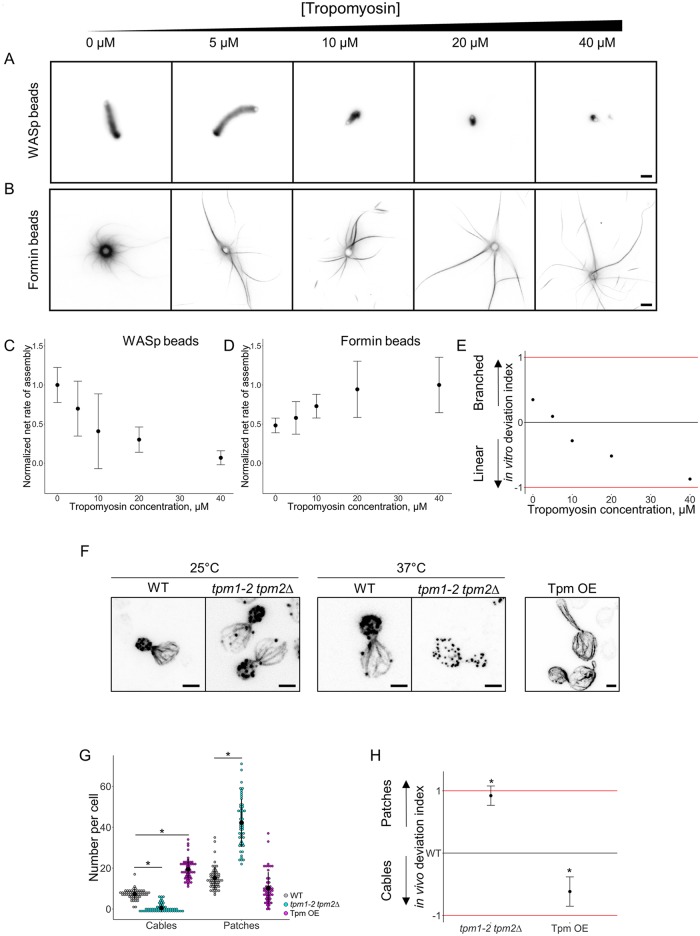
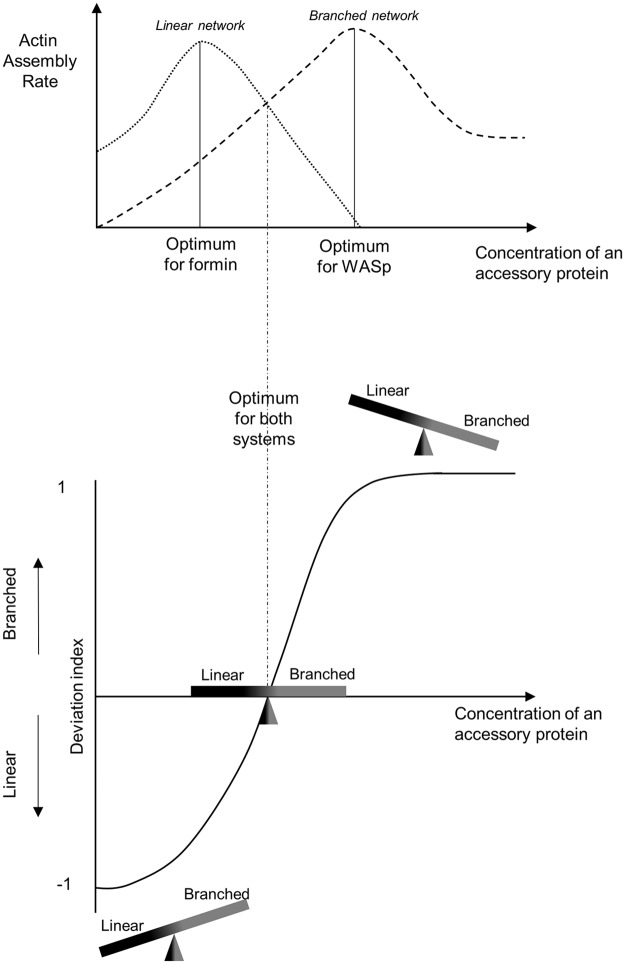
Within the cytoplasm of a single cell, several actin networks can coexist with distinct sizes, geometries, and protein compositions. These actin networks assemble in competition for a limited pool of proteins present in a common cellular environment. To predict how two distinct networks of actin filaments control this balance, the simultaneous assembly of actin-related protein 2/3 (Arp2/3)-branched networks and formin-linear networks of actin filaments around polystyrene microbeads was investigated with a range of actin accessory proteins (profilin, capping protein, actin-depolymerizing factor [ADF]/cofilin, and tropomyosin). Accessory proteins generally affected actin assembly rates for the distinct networks differently. These effects at the scale of individual actin networks were surprisingly not always correlated with corresponding loss-of-function phenotypes in cells. However, our observations agreed with a global interpretation, which compared relative actin assembly rates of individual actin networks. This work supports a general model in which the size of distinct actin networks is determined by their relative capacity to assemble in a common and competing environment.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
