

In cultured cells the baculovirus P10 protein forms two independent intracellular structures that play separate roles in occlusion body maturation and their release by nuclear disintegration
- PMID: 31181119
- PMCID: PMC6557513
- DOI: 10.1371/journal.ppat.1007827
In cultured cells the baculovirus P10 protein forms two independent intracellular structures that play separate roles in occlusion body maturation and their release by nuclear disintegration
Abstract
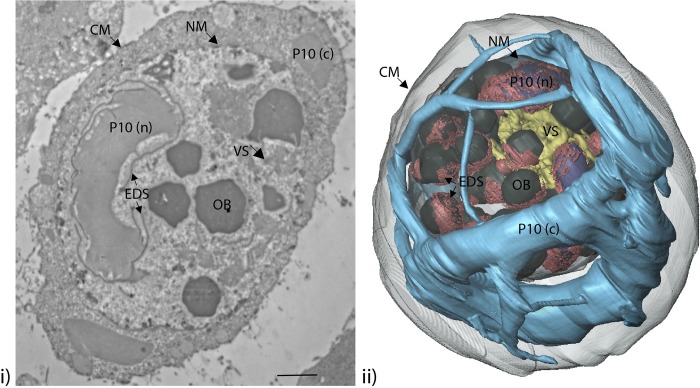
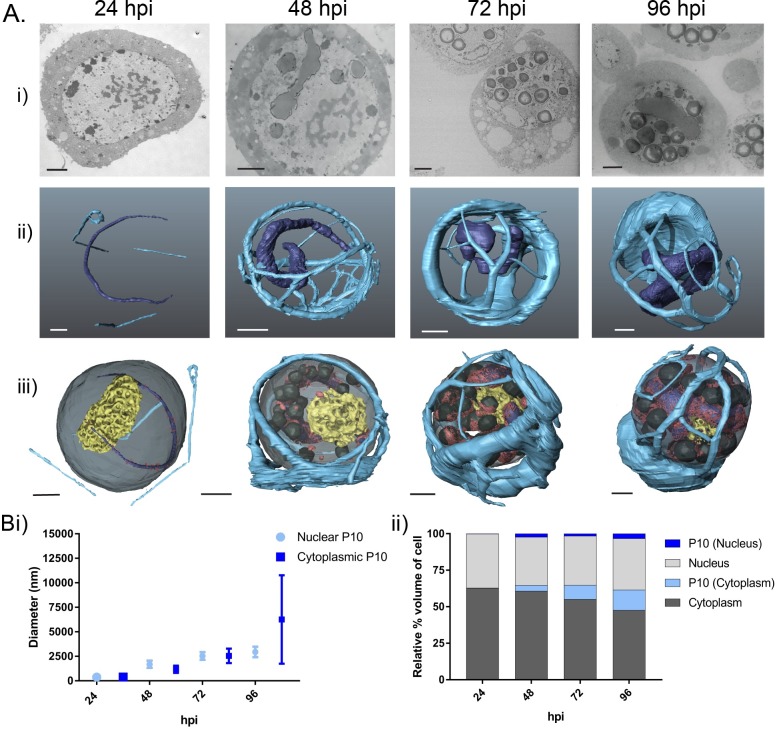
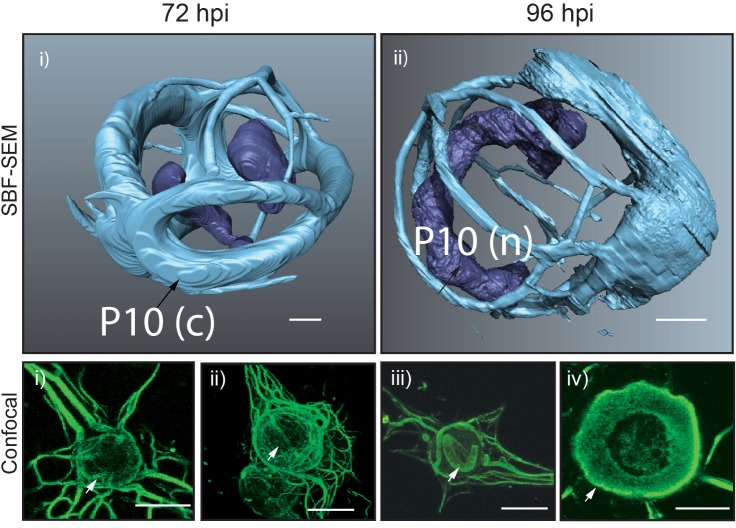
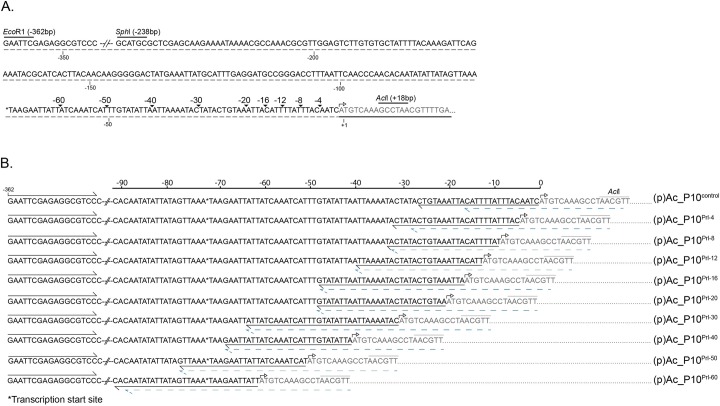
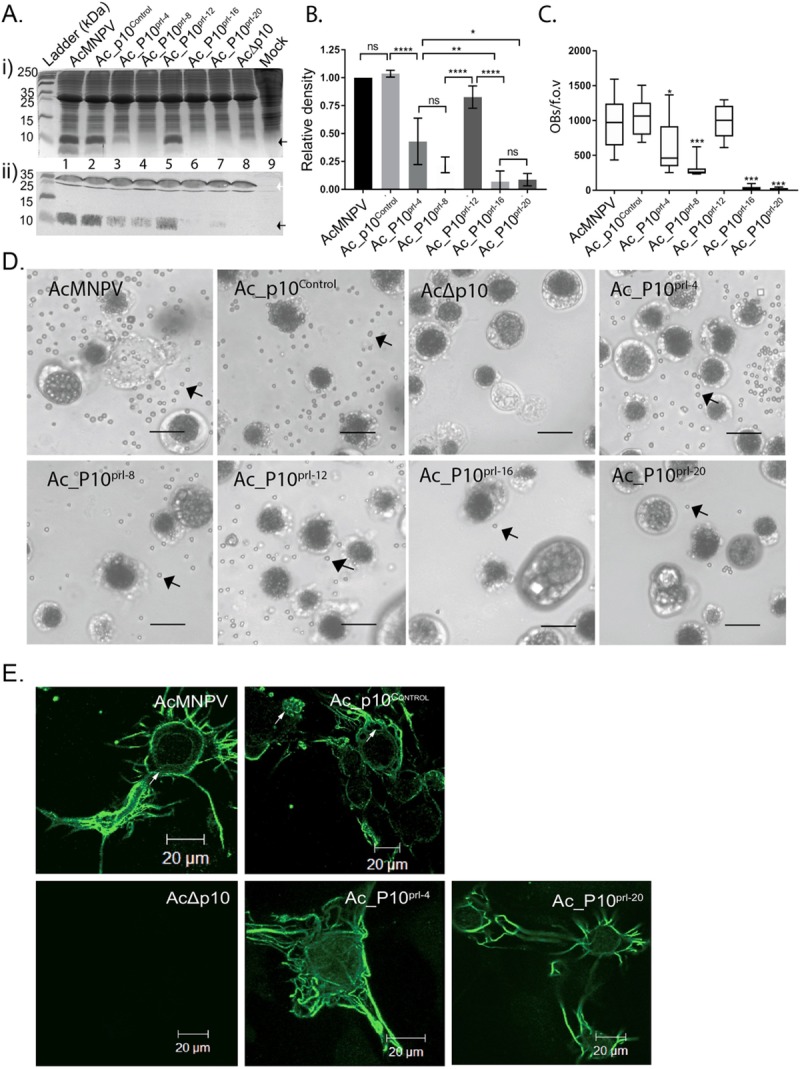
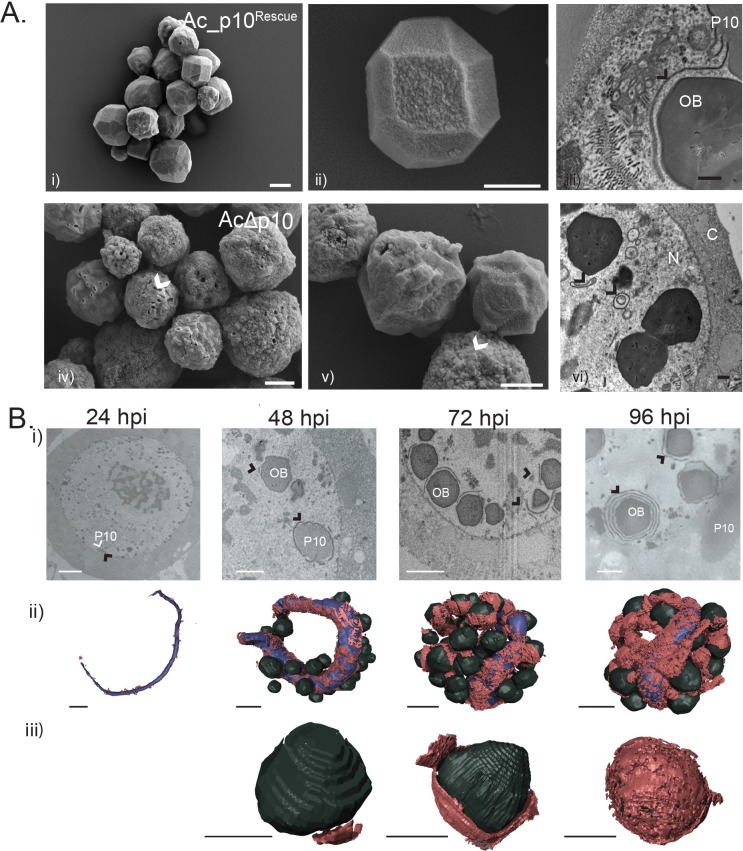
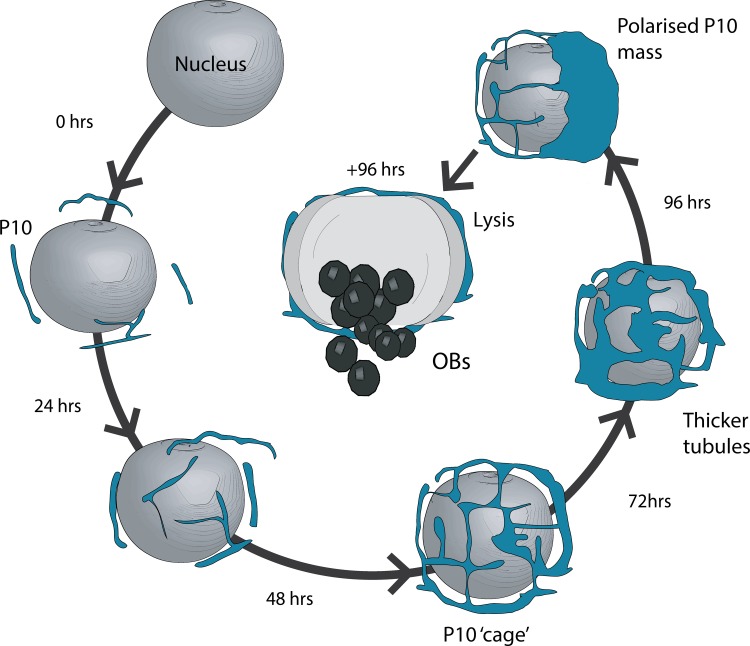
P10 is a small, abundant baculovirus protein that accumulates to high levels in the very late stages of the infection cycle. It is associated with a number of intracellular structures and implicated in diverse processes from occlusion body maturation to nuclear stability and lysis. However, studies have also shown that it is non-essential for virus replication, at least in cell culture. Here, we describe the use of serial block-face scanning electron microscopy to achieve high-resolution 3D characterisation of P10 structures within Trichoplusia ni TN-368 cells infected with Autographa californica multiple nucleopolyhedrovirus. This has enabled unparalleled visualisation of P10 and determined the independent formation of dynamic perinuclear and nuclear vermiform fibrous structures. Our 3D data confirm the sequence of ultrastructural changes that create a perinuclear cage from thin angular fibrils within the cytoplasm. Over the course of infection in cultured cells, the cage remodels to form a large polarised P10 mass and we suggest that these changes are critical for nuclear lysis to release occlusion bodies. In contrast, nuclear P10 forms a discrete vermiform structure that was observed in close spatial association with both electron dense spacers and occlusion bodies; supporting a previously suggested role for P10 and electron dense spacers in the maturation of occlusion bodies. We also demonstrate that P10 hyper-expression is critical for function. Decreasing levels of p10 expression, achieved by manipulation of promoter length, correlated with reduced P10 production, a lack of formation of P10 structures and a concomitant decrease in nuclear lysis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Phosphorylation Induces Structural Changes in the Autographa californica Nucleopolyhedrovirus P10 Protein.J Virol. 2017 Jun 9;91(13):e00002-17. doi: 10.1128/JVI.00002-17. Print 2017 Jul 1. J Virol. 2017. PMID: 28424279 Free PMC article.
-
The baculovirus P10 protein of Autographa californica nucleopolyhedrovirus forms two distinct cytoskeletal-like structures and associates with polyhedral occlusion bodies during infection.Virology. 2008 Feb 20;371(2):278-91. doi: 10.1016/j.virol.2007.09.043. Epub 2007 Nov 8. Virology. 2008. PMID: 17991504
-
Functional domains of the p10 protein of Autographa californica nuclear polyhedrosis virus.J Gen Virol. 1993 Apr;74 ( Pt 4):563-74. doi: 10.1099/0022-1317-74-4-563. J Gen Virol. 1993. PMID: 8468550
-
AcMNPV core gene ac109 is required for budded virion transport to the nucleus and for occlusion of viral progeny.PLoS One. 2012;7(9):e46146. doi: 10.1371/journal.pone.0046146. Epub 2012 Sep 26. PLoS One. 2012. PMID: 23049963 Free PMC article.
-
Specificity of baculovirus p10 functions.Virology. 1994 May 1;200(2):513-23. doi: 10.1006/viro.1994.1214. Virology. 1994. PMID: 8178439
Cited by
-
Dynamic chromatin accessibility profiling reveals changes in host genome organization in response to baculovirus infection.PLoS Pathog. 2020 Jun 8;16(6):e1008633. doi: 10.1371/journal.ppat.1008633. eCollection 2020 Jun. PLoS Pathog. 2020. PMID: 32511266 Free PMC article.
-
Sample preparation and data collection for serial block face scanning electron microscopy of mammalian cell monolayers.PLoS One. 2024 Aug 9;19(8):e0301284. doi: 10.1371/journal.pone.0301284. eCollection 2024. PLoS One. 2024. PMID: 39121154 Free PMC article.
-
Baculovirus entry into the central nervous system of Spodoptera exigua caterpillars is independent of the viral protein tyrosine phosphatase.Open Biol. 2024 Feb;14(2):230278. doi: 10.1098/rsob.230278. Epub 2024 Feb 21. Open Biol. 2024. PMID: 38378139 Free PMC article.
-
BV/ODV-E26 is a conserved baculoviral inhibitory factor for optimizing viral virulence in lepidopteran hosts.iScience. 2024 Dec 31;28(2):111723. doi: 10.1016/j.isci.2024.111723. eCollection 2025 Feb 21. iScience. 2024. PMID: 39898022 Free PMC article.
References
-
- Pearson MN, Russell RL, Rohrmann GF, Beaudreau GS. p39, a major baculovirus structural protein: immunocytochemical characterization and genetic location. Virology. 1988;167(2):407–13. - PubMed
-
- Boucias DG, Pendland JC. Baculoviruses In: Boucias DG, Pendland JC, editors. Principles of Insect Pathology; Norwell, New York: Kluwer Academic Publishers; 1998. p. 111–46.
-
- Summers MD, Arnott HJ. Ultrastructural studies on inclusion formation and virus occlusion in nuclear polyhedrosis and granulosis virus-infected cells of Trichoplusia ni (Hubner). J Ultrastruct Res. 1969;28(5):462–80. . - PubMed
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
