

Neuro-genetic plasticity of Caenorhabditis elegans behavioral thermal tolerance
- PMID: 31182018
- PMCID: PMC6558720
- DOI: 10.1186/s12868-019-0510-z
Neuro-genetic plasticity of Caenorhabditis elegans behavioral thermal tolerance
Abstract
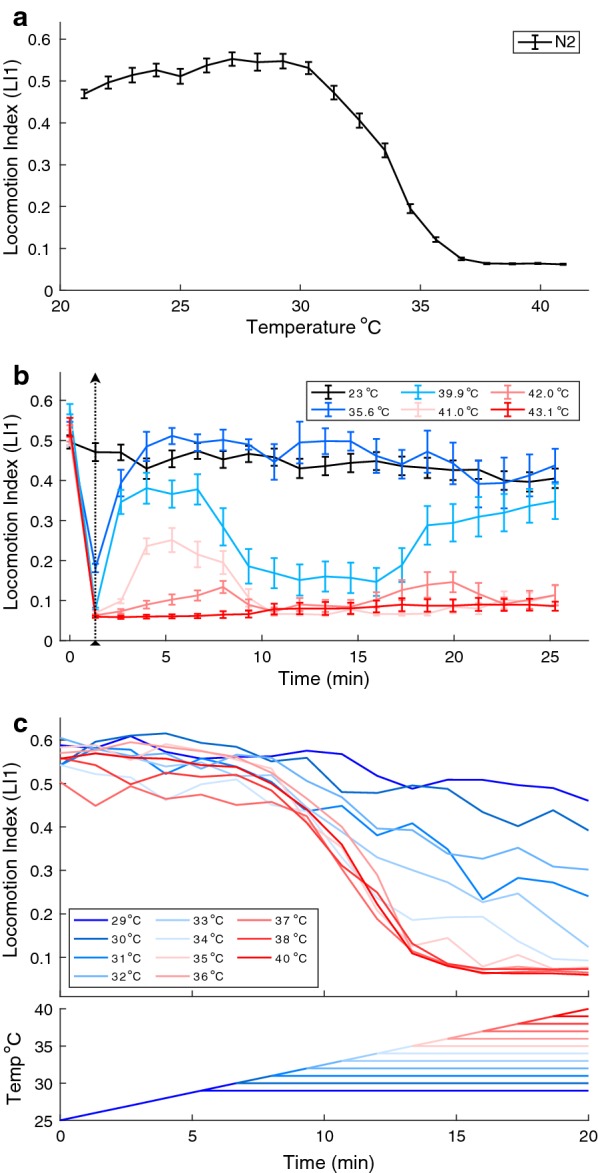
Background: Animal responses to thermal stimuli involve intricate contributions of genetics, neurobiology and physiology, with temperature variation providing a pervasive environmental factor for natural selection. Thermal behavior thus exemplifies a dynamic trait that requires non-trivial phenotypic summaries to appropriately capture the trait in response to a changing environment. To characterize the deterministic and plastic components of thermal responses, we developed a novel micro-droplet assay of nematode behavior that permits information-dense summaries of dynamic behavioral phenotypes as reaction norms in response to increasing temperature (thermal tolerance curves, TTC).
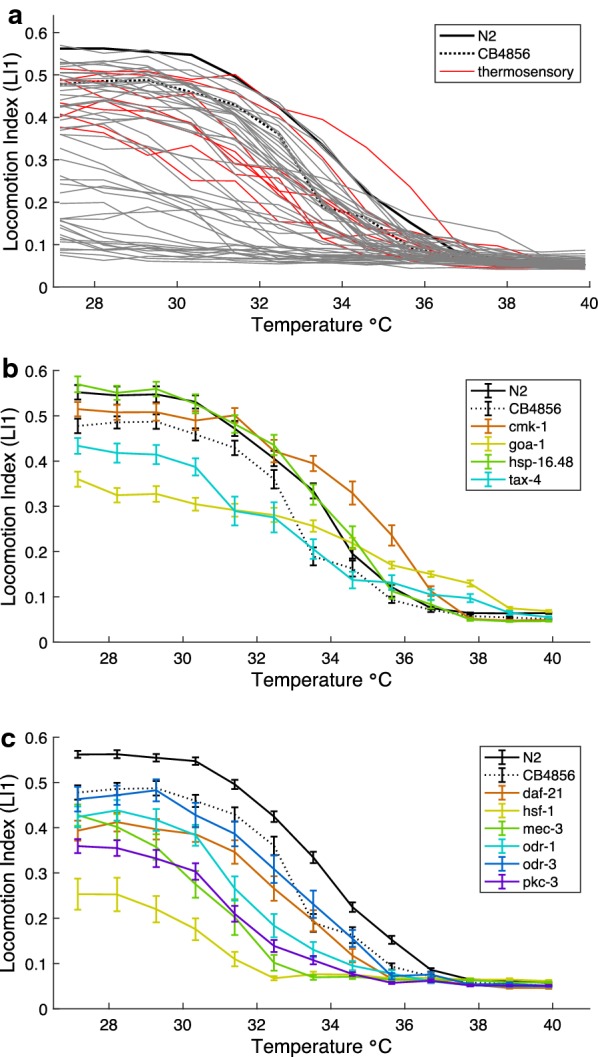
Results: We found that C. elegans TTCs shift predictably with rearing conditions and developmental stage, with significant differences between distinct wildtype genetic backgrounds. Moreover, after screening TTCs for 58 C. elegans genetic mutant strains, we determined that genes affecting thermosensation, including cmk-1 and tax-4, potentially play important roles in the behavioral control of locomotion at high temperature, implicating neural decision-making in TTC shape rather than just generalized physiological limits. However, expression of the transient receptor potential ion channel TRPA-1 in the nervous system is not sufficient to rescue rearing-dependent plasticity in TTCs conferred by normal expression of this gene, indicating instead a role for intestinal signaling involving TRPA-1 in the adaptive plasticity of thermal performance.
Conclusions: These results implicate nervous system and non-nervous system contributions to behavior, in addition to basic cellular physiology, as key mediators of evolutionary responses to selection from temperature variation in nature.
Keywords: C. elegans behavior; Computational ethology; Thermal ecology.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
-
- Fuller JL. Behavior genetics. Annu Rev Psychol. 1960;11(1):41–70. doi: 10.1146/annurev.ps.11.020160.000353. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
