

Shear stress regulation of miR-93 and miR-484 maturation through nucleolin
- PMID: 31182601
- PMCID: PMC6600934
- DOI: 10.1073/pnas.1902844116
Shear stress regulation of miR-93 and miR-484 maturation through nucleolin
Abstract
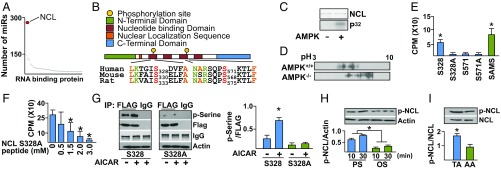
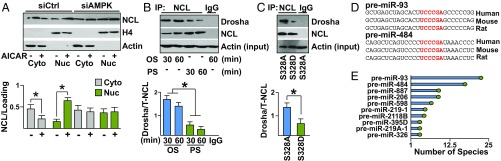
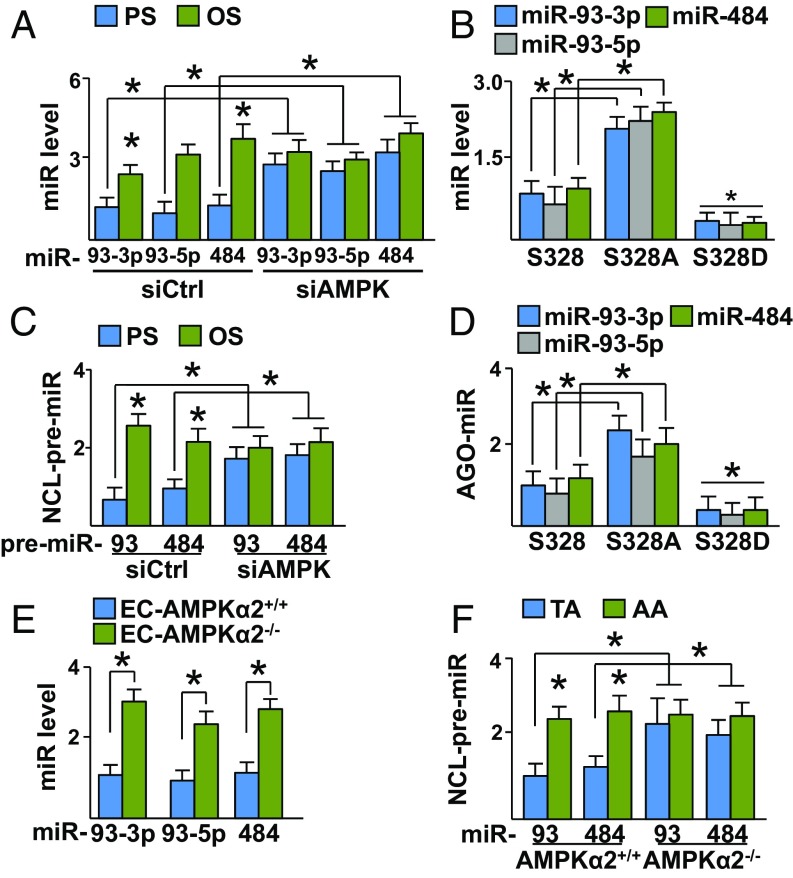
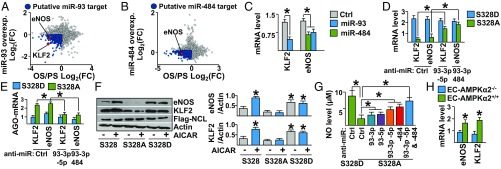
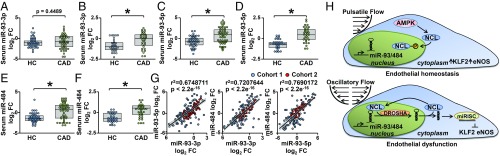
Pulsatile shear (PS) and oscillatory shear (OS) elicit distinct mechanotransduction signals that maintain endothelial homeostasis or induce endothelial dysfunction, respectively. A subset of microRNAs (miRs) in vascular endothelial cells (ECs) are differentially regulated by PS and OS, but the regulation of the miR processing and its implications in EC biology by shear stress are poorly understood. From a systematic in silico analysis for RNA binding proteins that regulate miR processing, we found that nucleolin (NCL) is a major regulator of miR processing in response to OS and essential for the maturation of miR-93 and miR-484 that target mRNAs encoding Krüppel-like factor 2 (KLF2) and endothelial nitric oxide synthase (eNOS). Additionally, anti-miR-93 and anti-miR-484 restore KLF2 and eNOS expression and NO bioavailability in ECs under OS. Analysis of posttranslational modifications of NCL identified that serine 328 (S328) phosphorylation by AMP-activated protein kinase (AMPK) was a major PS-activated event. AMPK phosphorylation of NCL sequesters it in the nucleus, thereby inhibiting miR-93 and miR-484 processing and their subsequent targeting of KLF2 and eNOS mRNA. Elevated levels of miR-93 and miR-484 were found in sera collected from individuals afflicted with coronary artery disease in two cohorts. These findings provide translational relevance of the AMPK-NCL-miR-93/miR-484 axis in miRNA processing in EC health and coronary artery disease.
Keywords: AMPK; endothelial cells; miRNA; nucleolin; shear stress.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Demolli S., et al. , Shear stress-regulated miR-27b controls pericyte recruitment by repressing SEMA6A and SEMA6D. Cardiovasc. Res. 113, 681–691 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
