

Rho kinase-dependent apical constriction counteracts M-phase apical expansion to enable mouse neural tube closure
- PMID: 31182644
- PMCID: PMC6633395
- DOI: 10.1242/jcs.230300
Rho kinase-dependent apical constriction counteracts M-phase apical expansion to enable mouse neural tube closure
Abstract
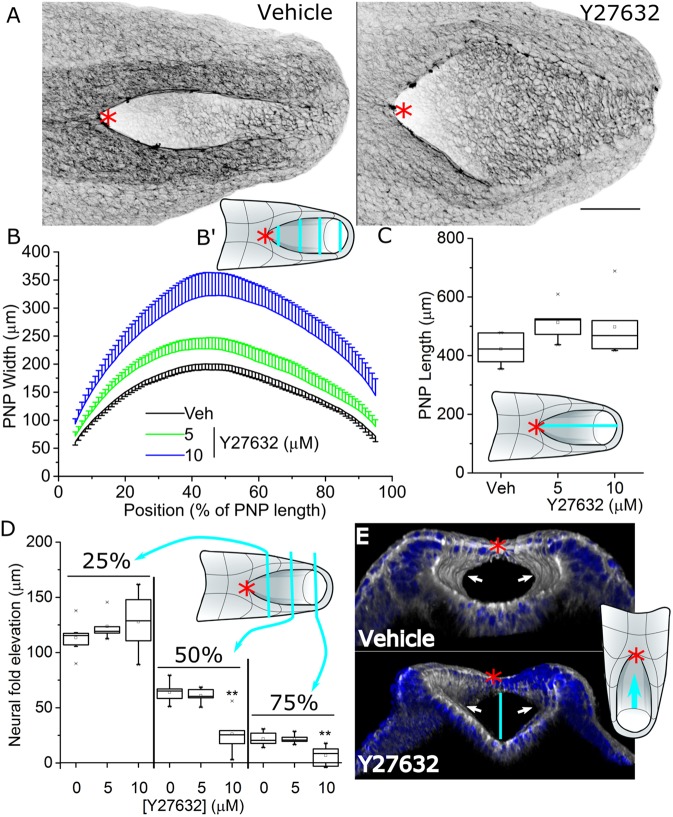
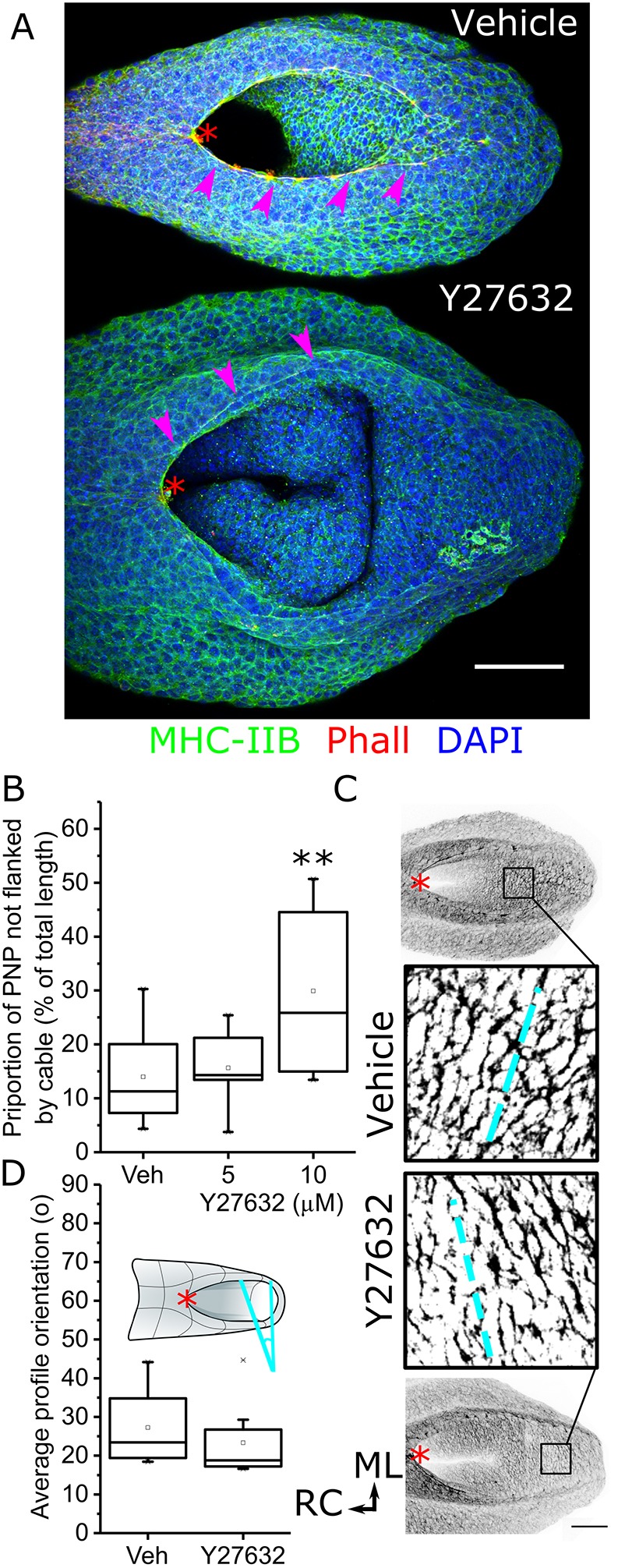
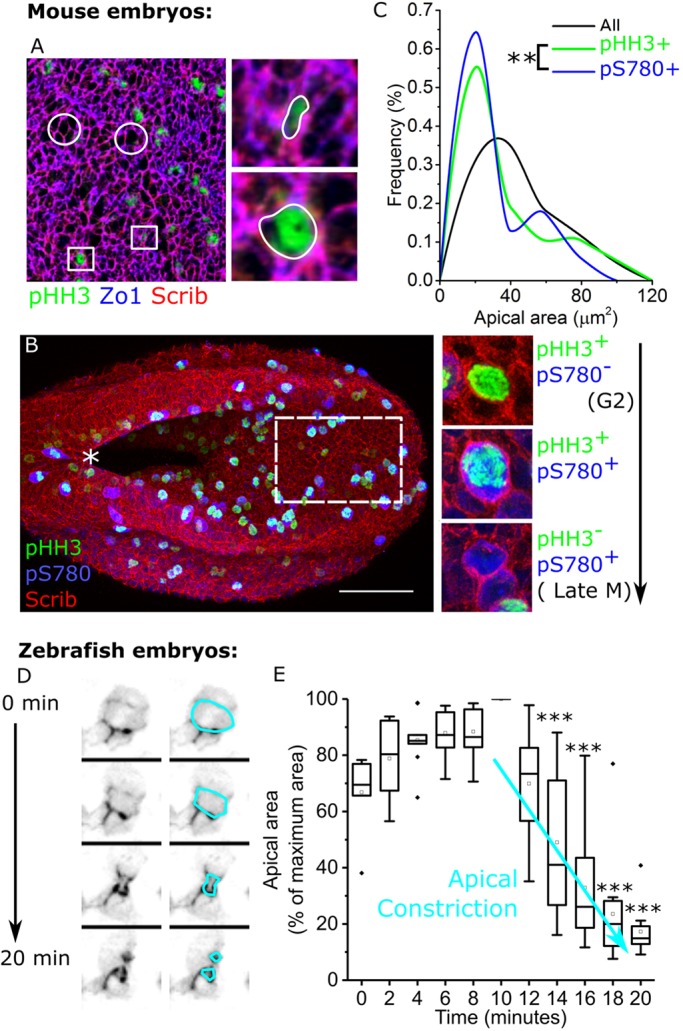
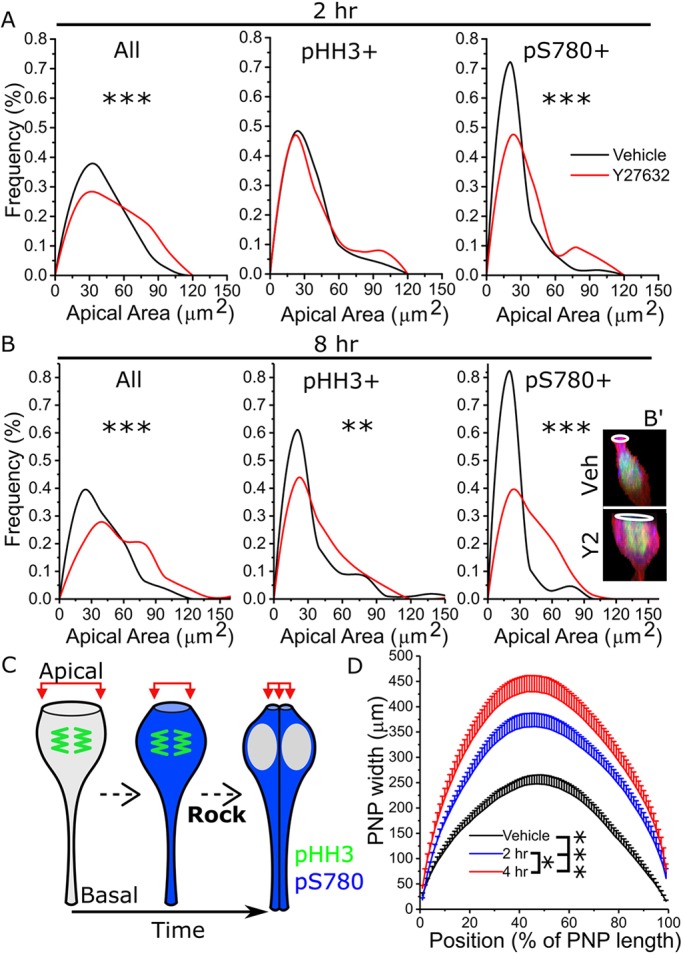
Cellular generation of mechanical forces required to close the presumptive spinal neural tube, the 'posterior neuropore' (PNP), involves interkinetic nuclear migration (INM) and apical constriction. Both processes change the apical surface area of neuroepithelial cells, but how they are biomechanically integrated is unknown. Rho kinase (Rock; herein referring to both ROCK1 and ROCK2) inhibition in mouse whole embryo culture progressively widens the PNP. PNP widening is not caused by increased mechanical tension opposing closure, as evidenced by diminished recoil following laser ablation. Rather, Rock inhibition diminishes neuroepithelial apical constriction, producing increased apical areas in neuroepithelial cells despite diminished tension. Neuroepithelial apices are also dynamically related to INM progression, with the smallest dimensions achieved in cells positive for the pan-M phase marker Rb phosphorylated at S780 (pRB-S780). A brief (2 h) Rock inhibition selectively increases the apical area of pRB-S780-positive cells, but not pre-anaphase cells positive for phosphorylated histone 3 (pHH3+). Longer inhibition (8 h, more than one cell cycle) increases apical areas in pHH3+ cells, suggesting cell cycle-dependent accumulation of cells with larger apical surfaces during PNP widening. Consequently, arresting cell cycle progression with hydroxyurea prevents PNP widening following Rock inhibition. Thus, Rock-dependent apical constriction compensates for the PNP-widening effects of INM to enable progression of closure.This article has an associated First Person interview with the first authors of the paper.
Keywords: Apical constriction; Biomechanics; F-actin; Interkinetic nuclear migration; Posterior neuropore; Rock.
© 2019. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsA.J.C. acts as a paid consultant for ViiV Healthcare Limited, with fees going to support his research programme. No other competing interests are declared.
Figures



References
-
- Bush K. T., Lynch F. J., DeNittis A. S., Steinberg A. B., Lee H.-Y. and Nagele R. G. (1990). Neural tube formation in the mouse: a morphometric and computerized three-dimensional reconstruction study of the relationship between apical constriction of neuroepithelial cells and the shape of the neuroepithelium. Anat. Embryol. 181, 49-58. 10.1007/BF00189727 - DOI - PubMed
-
- Cavadino A., Prieto-Merino D., Addor M.-C., Arriola L., Bianchi F., Draper E., Garne E., Greenlees R., Haeusler M., Khoshnood B. et al. (2016). Use of hierarchical models to analyze European trends in congenital anomaly prevalence. Birth Defects Res. A Clin. Mol. Teratol. 106, 480-488. 10.1002/bdra.23515 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
