

The roles of structural dynamics in the cellular functions of RNAs
- PMID: 31182864
- PMCID: PMC7656661
- DOI: 10.1038/s41580-019-0136-0
The roles of structural dynamics in the cellular functions of RNAs
Abstract
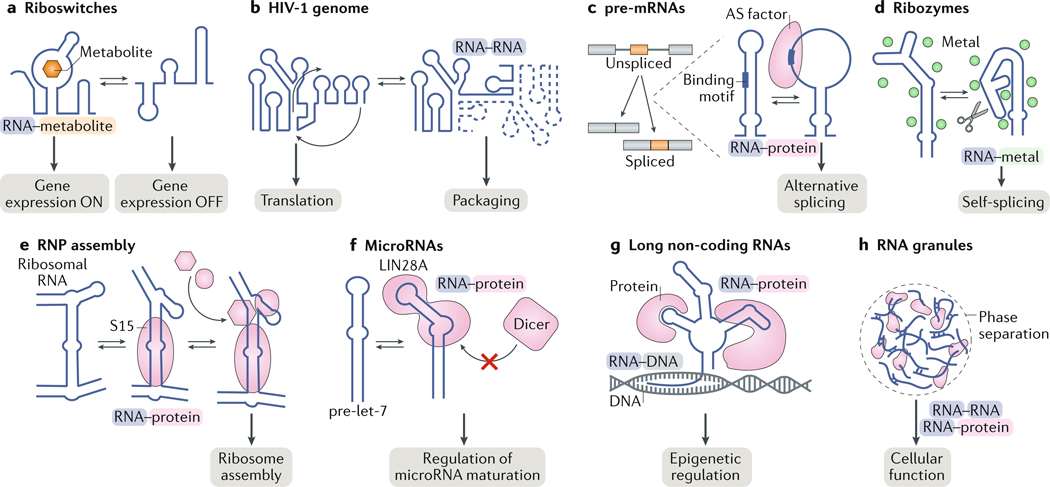
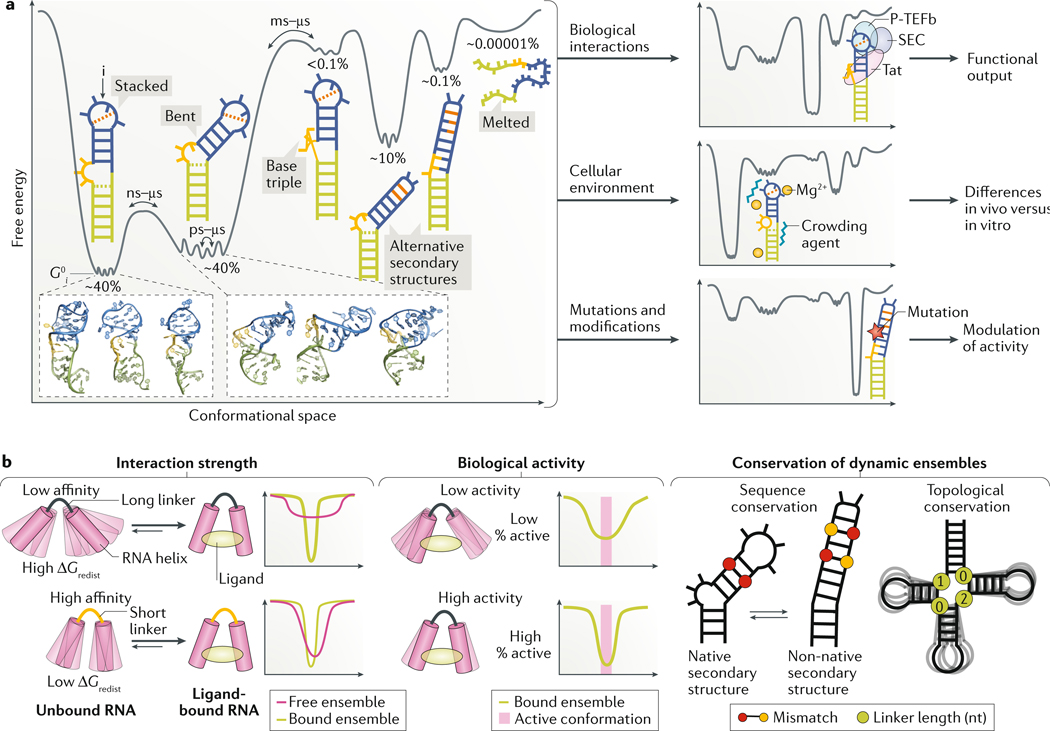
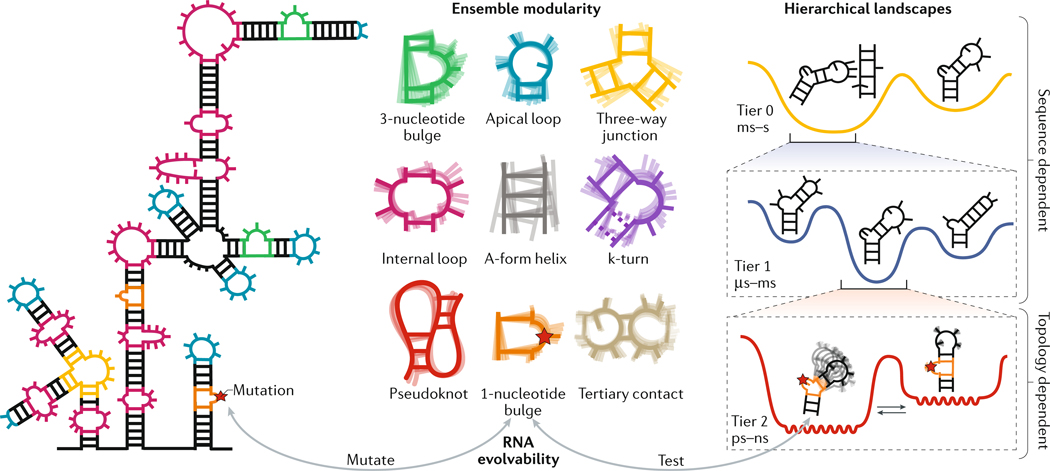
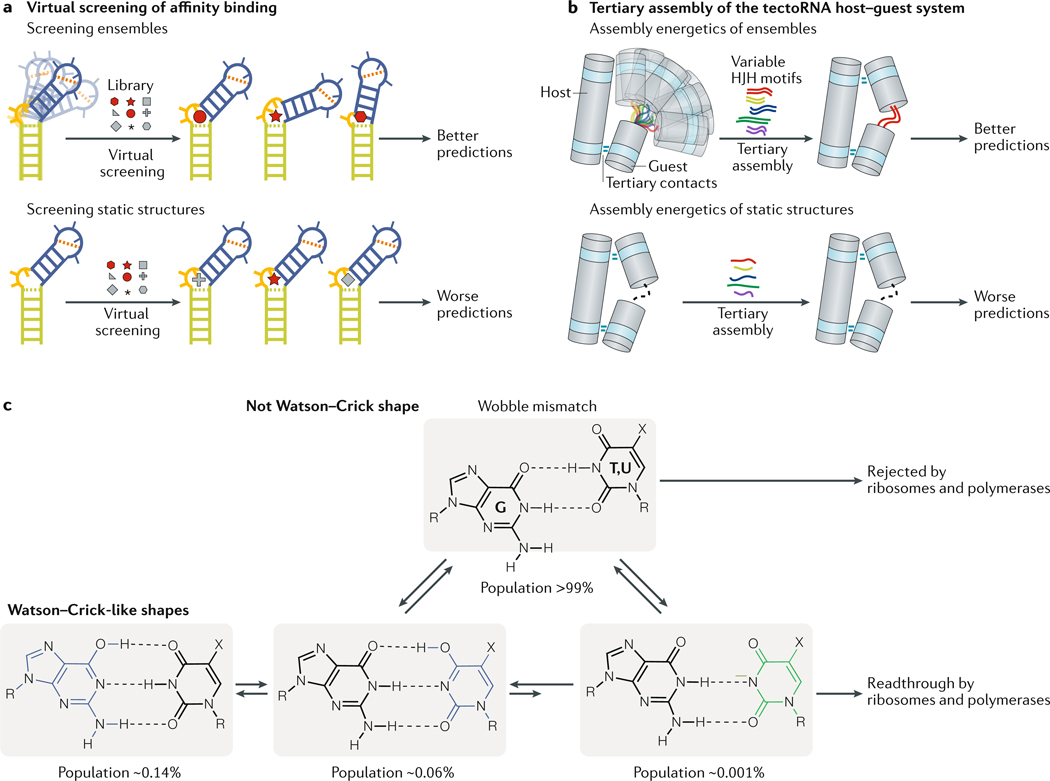
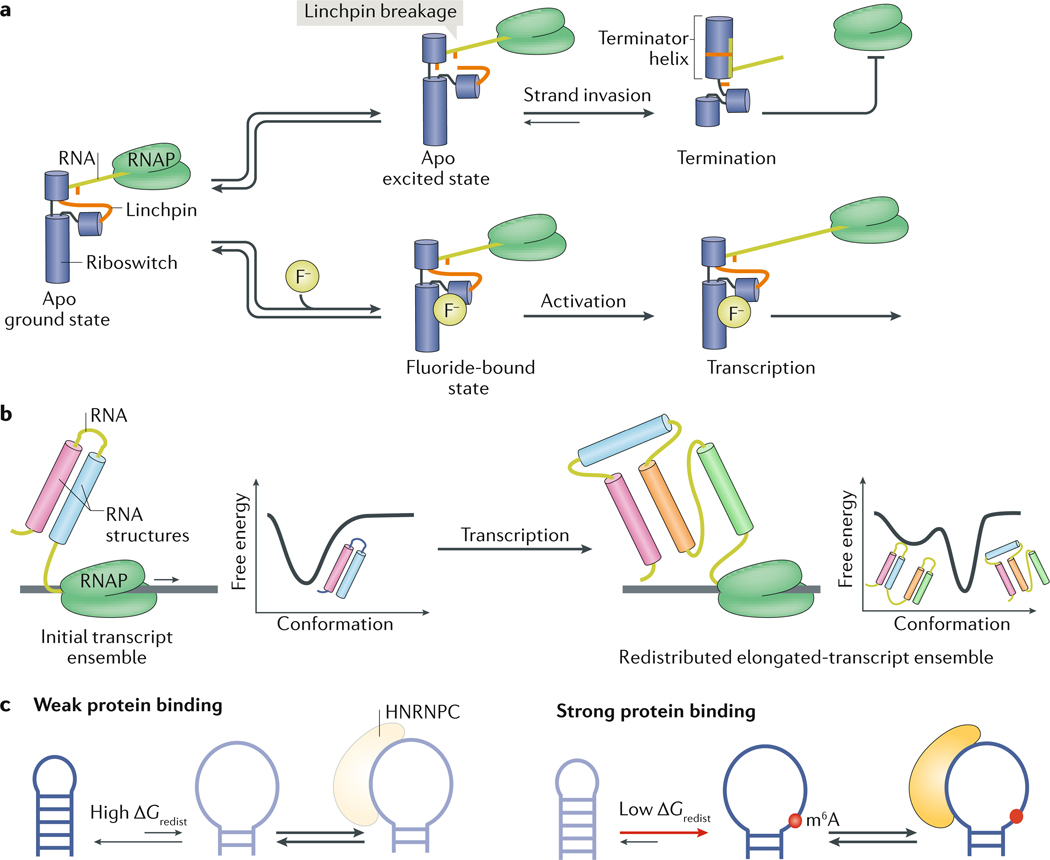
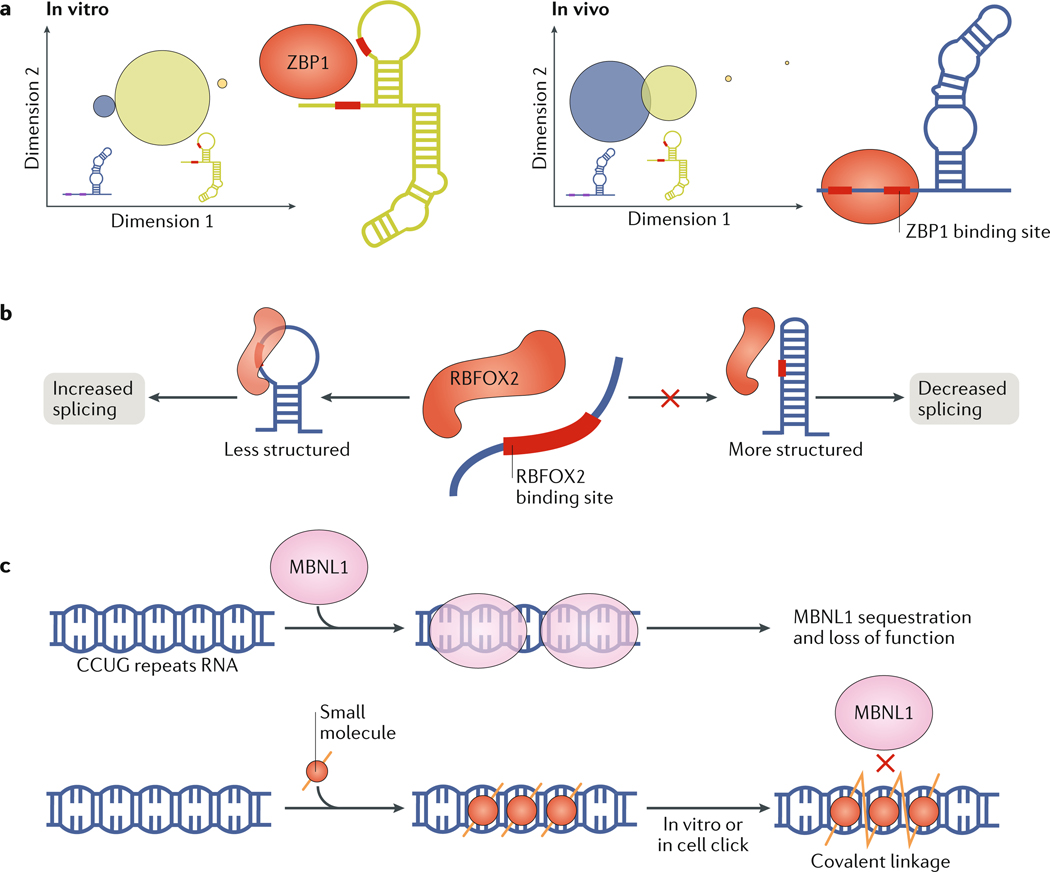
RNAs fold into 3D structures that range from simple helical elements to complex tertiary structures and quaternary ribonucleoprotein assemblies. The functions of many regulatory RNAs depend on how their 3D structure changes in response to a diverse array of cellular conditions. In this Review, we examine how the structural characterization of RNA as dynamic ensembles of conformations, which form with different probabilities and at different timescales, is improving our understanding of RNA function in cells. We discuss the mechanisms of gene regulation by microRNAs, riboswitches, ribozymes, post-transcriptional RNA modifications and RNA-binding proteins, and how the cellular environment and processes such as liquid-liquid phase separation may affect RNA folding and activity. The emerging RNA-ensemble-function paradigm is changing our perspective and understanding of RNA regulation, from in vitro to in vivo and from descriptive to predictive.
Figures
References
-
- Weber G. Energetics of ligand binding to proteins. Adv. Protein Chem 29, 1–83 (1975). - PubMed
-
- Cech TR & Steitz JA The noncoding RNA revolution-trashing old rules to forge new ones. Cell 157, 77–94 (2014). - PubMed
-
- Esteller M. Non-coding RNAs in human disease. Nat. Rev. Genet 12, 861–874 (2011). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
