

Evidence for Similar Prefrontal Structural and Functional Alterations in Male and Female Rats Following Chronic Stress or Glucocorticoid Exposure
- PMID: 31184364
- PMCID: PMC7029687
- DOI: 10.1093/cercor/bhz092
Evidence for Similar Prefrontal Structural and Functional Alterations in Male and Female Rats Following Chronic Stress or Glucocorticoid Exposure
Abstract
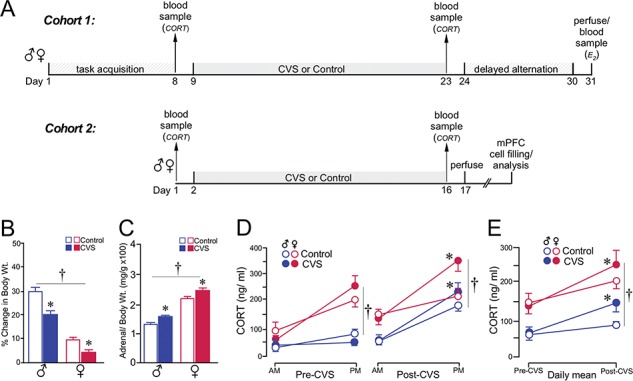
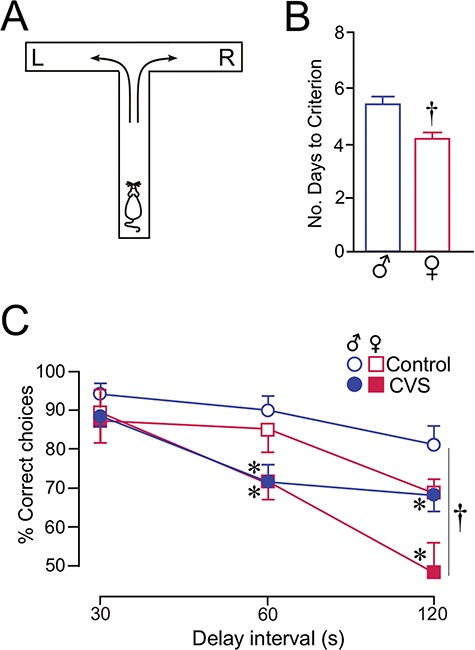
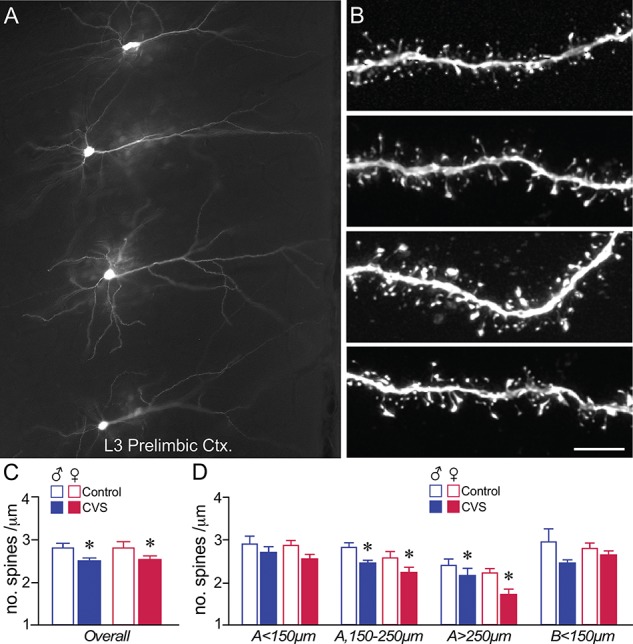
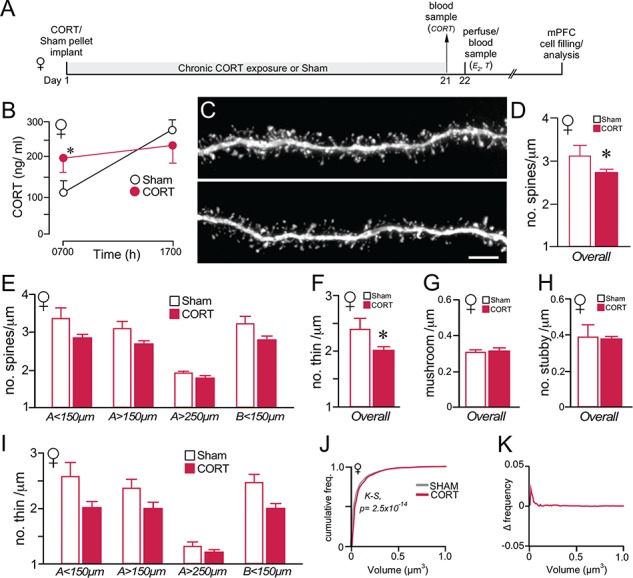
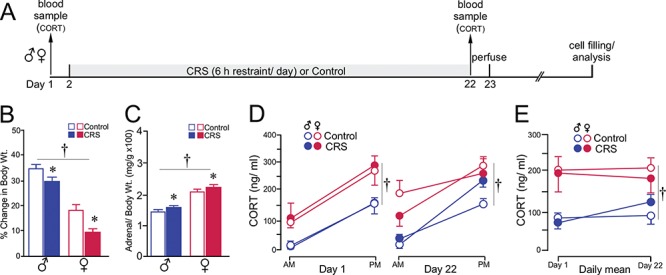
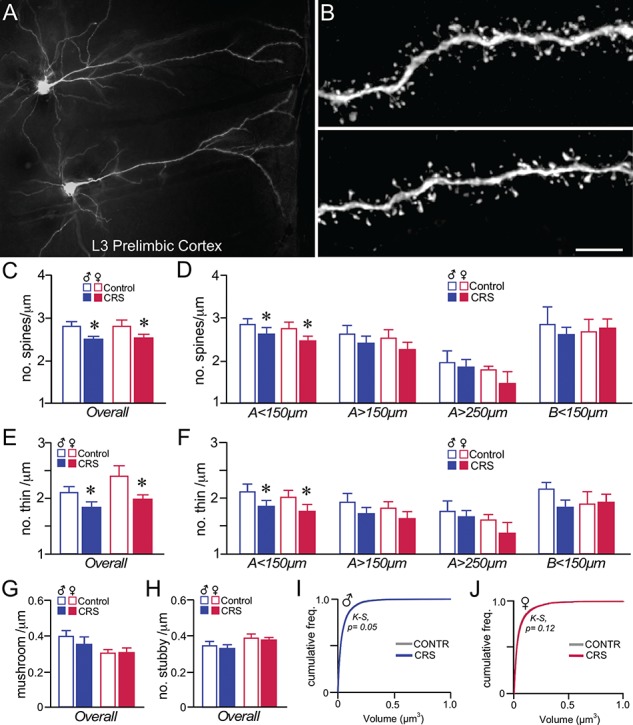
Previous work of ours and others has documented regressive changes in neuronal architecture and function in the medial prefrontal cortex (mPFC) of male rats following chronic stress. As recent focus has shifted toward understanding whether chronic stress effects on mPFC are sexually dimorphic, here we undertake a comprehensive analysis to address this issue. First, we show that chronic variable stress (14-day daily exposure to different challenges) resulted in a comparable degree of adrenocortical hyperactivity, working memory impairment, and dendritic spine loss in mPFC pyramidal neurons in both sexes. Next, exposure of female rats to 21-day regimen of corticosterone resulted in a similar pattern of mPFC dendritic spine attrition and increase in spine volume. Finally, we examined the effects of another widely used regimen, chronic restraint stress (CRS, 21-day of daily 6-h restraint), on dendritic spine changes in mPFC in both sexes. CRS resulted in response decrements in adrenocortical output (habituation), and induced a pattern of consistent, but less widespread, dendritic spine loss similar to the foregoing challenges. Our data suggest that chronic stress or glucocorticoid exposure induces a relatively undifferentiated pattern of structural and functional alterations in mPFC in both males and females.
Keywords: corticosterone; delayed alternation; dendritic spine; prelimbic; sex differences.
© The Author(s) 2019. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures

References
-
- Arnsten AF. 2000. Stress impairs prefrontal cortical function in rats and monkeys: role of dopamine D1 and norepinephrine a-1 receptor mechanisms. Progress in Brain Research. 126:183–192. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
