

Genome-wide RNA pol II initiation and pausing in neural progenitors of the rat
- PMID: 31185909
- PMCID: PMC6558777
- DOI: 10.1186/s12864-019-5829-4
Genome-wide RNA pol II initiation and pausing in neural progenitors of the rat
Abstract
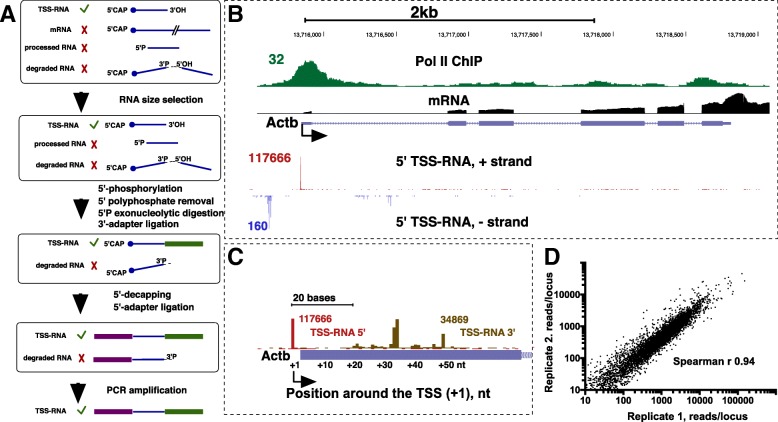
Background: Global RNA sequencing technologies have revealed widespread RNA polymerase II (Pol II) transcription outside of gene promoters. Small 5'-capped RNA sequencing (Start-seq) originally developed for the detection of promoter-proximal Pol II pausing has helped improve annotation of Transcription Start Sites (TSSs) of genes as well as identification of non-genic regulatory elements. However, apart from the most well studied genomes of human and mouse, mammalian transcription has not been profiled with sufficiently high precision.
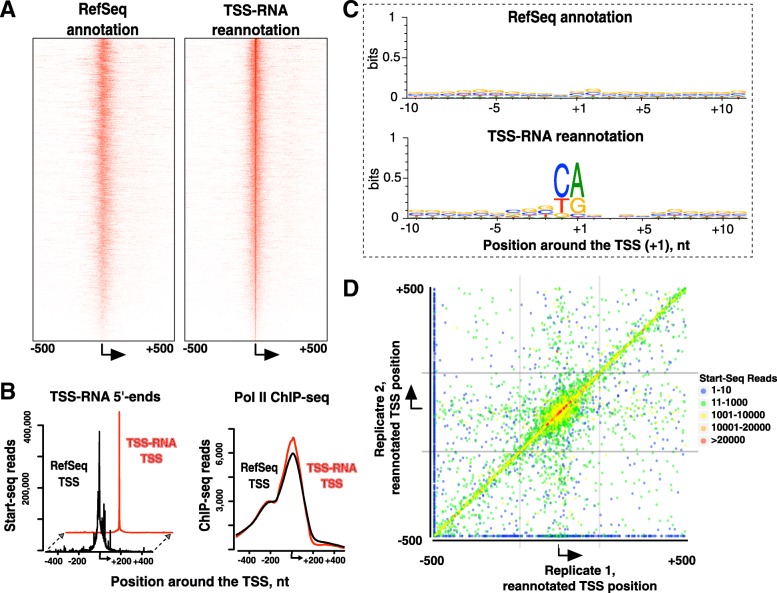
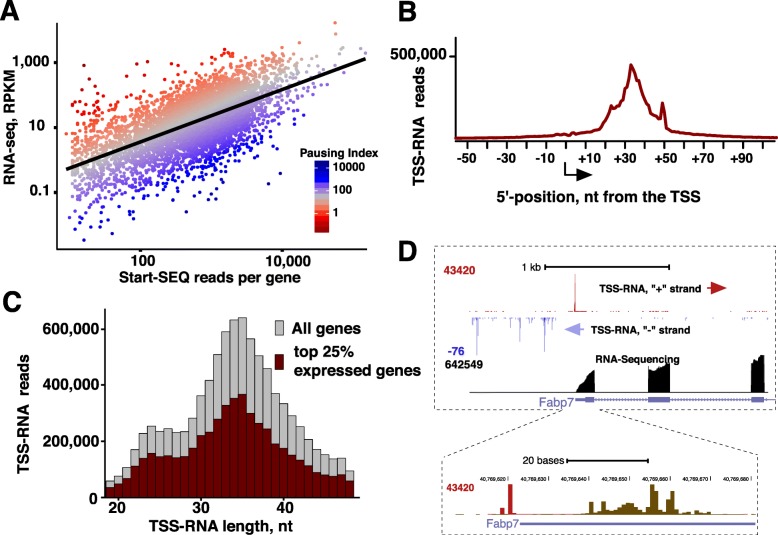
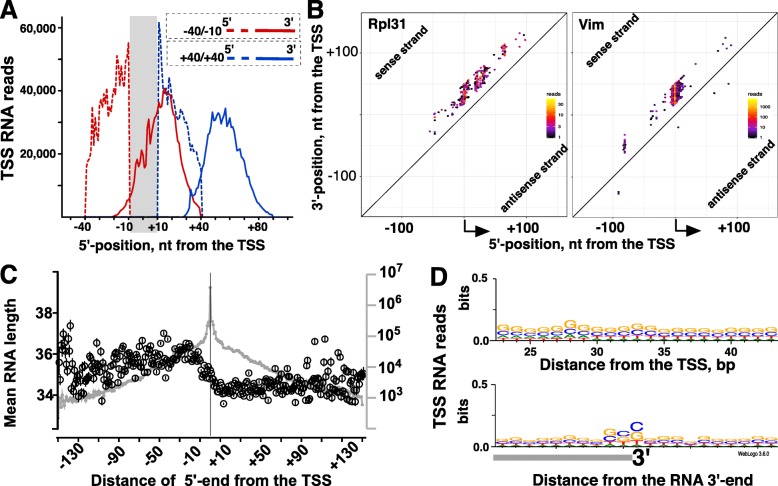
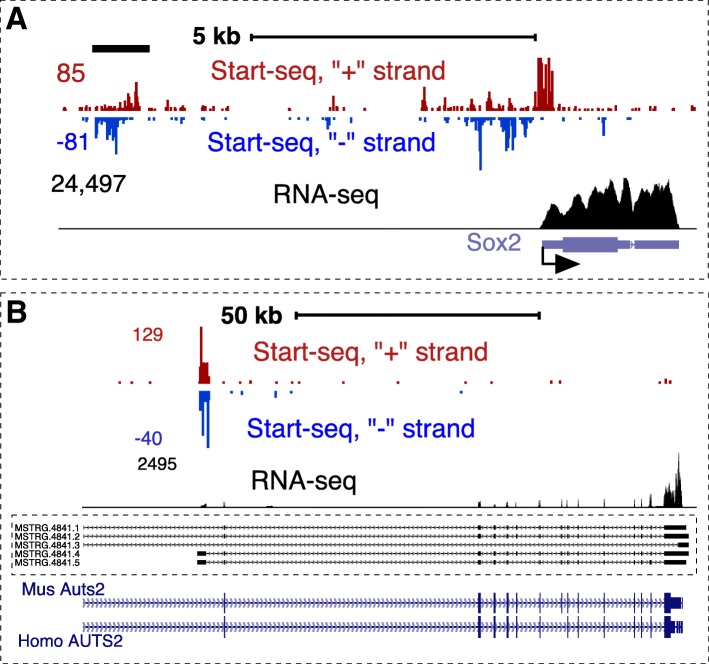
Results: We prepared and sequenced Start-seq libraries from rat (Rattus norgevicus) primary neural progenitor cells. Over 48 million uniquely mappable reads from two independent biological replicates allowed us to define the TSSs of 7365 known genes in the rn6 genome, reannotating 2503 TSSs by more than 5 base pairs, characterize promoter-associated antisense transcription, and profile Pol II pausing. By combining TSS data with polyA-selected RNA sequencing, we also identified thousands of potential new genes producing stable RNA as well as non-genic transcripts representing possible regulatory elements.
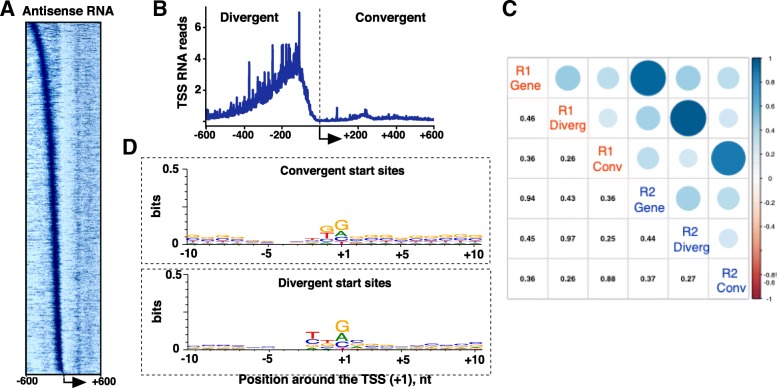
Conclusions: Our study has produced the first Start-seq dataset for the rat. Apart from profiling transcription initiation, our data reaffirm the prevalence of Pol II pausing across the rat genome and indicate conservation of pausing mechanisms across metazoan genomes. We suggest that pausing location, at least in mammals, is constrained by a distance from initiation of transcription, whether it occurs at or outside of a gene promoter. Abundant antisense transcription initiation around protein coding genes indicates that Pol II recruited to the vicinity of a promoter is distributed to available start sites of transcription at either DNA strand. Transcriptome profiling of neural progenitors presented here will facilitate further studies of other rat cell types as well as other organisms.
Keywords: Promoters; RNA pol II; Small RNA; Transcription.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Ssl2/TFIIH function in transcription start site scanning by RNA polymerase II in Saccharomyces cerevisiae.Elife. 2021 Oct 15;10:e71013. doi: 10.7554/eLife.71013. Elife. 2021. PMID: 34652274 Free PMC article.
-
RNA Polymerase II Promoter-Proximal Pausing and Release to Elongation Are Key Steps Regulating Herpes Simplex Virus 1 Transcription.J Virol. 2020 Feb 14;94(5):e02035-19. doi: 10.1128/JVI.02035-19. Print 2020 Feb 14. J Virol. 2020. PMID: 31826988 Free PMC article.
-
Pausing sites of RNA polymerase II on actively transcribed genes are enriched in DNA double-stranded breaks.J Biol Chem. 2020 Mar 20;295(12):3990-4000. doi: 10.1074/jbc.RA119.011665. Epub 2020 Feb 6. J Biol Chem. 2020. PMID: 32029477 Free PMC article.
-
Use of the nuclear walk-on methodology to determine sites of RNA polymerase II initiation and pausing and quantify nascent RNAs in cells.Methods. 2019 Apr 15;159-160:165-176. doi: 10.1016/j.ymeth.2019.02.003. Epub 2019 Feb 8. Methods. 2019. PMID: 30743000 Free PMC article. Review.
-
RNA polymerase II pausing in development: orchestrating transcription.Open Biol. 2022 Jan;12(1):210220. doi: 10.1098/rsob.210220. Epub 2022 Jan 5. Open Biol. 2022. PMID: 34982944 Free PMC article. Review.
Cited by
-
Cis-Regulatory Elements in Mammals.Int J Mol Sci. 2023 Dec 26;25(1):343. doi: 10.3390/ijms25010343. Int J Mol Sci. 2023. PMID: 38203513 Free PMC article. Review.
-
ERK2-topoisomerase II regulatory axis is important for gene activation in immediate early genes.Nat Commun. 2023 Dec 14;14(1):8341. doi: 10.1038/s41467-023-44089-y. Nat Commun. 2023. PMID: 38097570 Free PMC article.
-
Global approaches for profiling transcription initiation.Cell Rep Methods. 2021 Sep 27;1(5):100081. doi: 10.1016/j.crmeth.2021.100081. Epub 2021 Sep 16. Cell Rep Methods. 2021. PMID: 34632443 Free PMC article. Review.
-
Disrupting the RNA polymerase II transcription cycle through CDK7 inhibition ameliorates inflammatory arthritis.Sci Transl Med. 2024 Nov 20;16(774):eadq5091. doi: 10.1126/scitranslmed.adq5091. Epub 2024 Nov 20. Sci Transl Med. 2024. PMID: 39565872
-
Proteasome Inhibition Reprograms Chromatin Landscape in Breast Cancer.Cancer Res Commun. 2024 Apr 16;4(4):1082-1099. doi: 10.1158/2767-9764.CRC-23-0476. Cancer Res Commun. 2024. PMID: 38625038 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
