

Cardiolipin-induced activation of pyruvate dehydrogenase links mitochondrial lipid biosynthesis to TCA cycle function
- PMID: 31186346
- PMCID: PMC6663869
- DOI: 10.1074/jbc.RA119.009037
Cardiolipin-induced activation of pyruvate dehydrogenase links mitochondrial lipid biosynthesis to TCA cycle function
Abstract
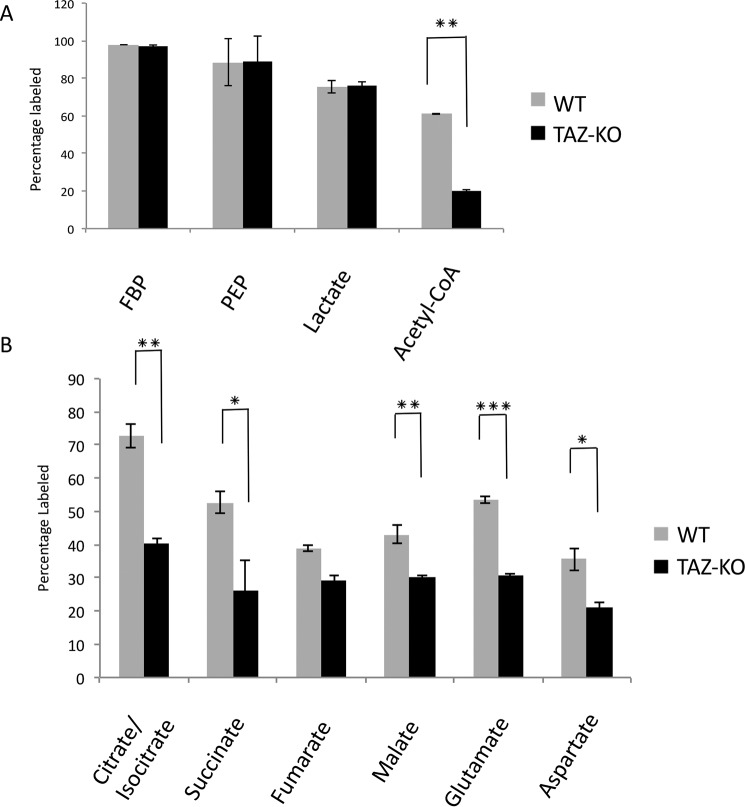
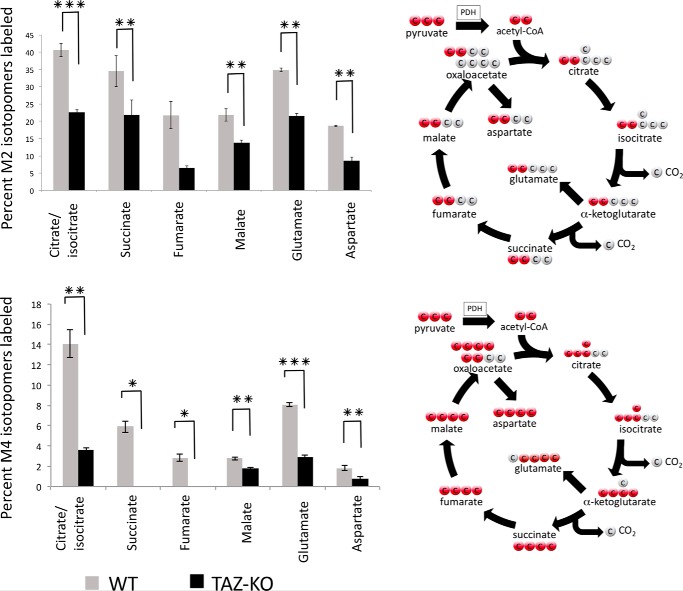
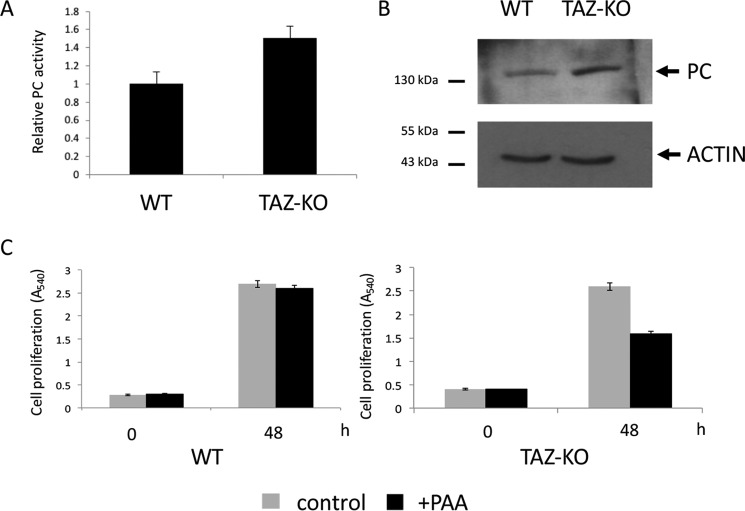
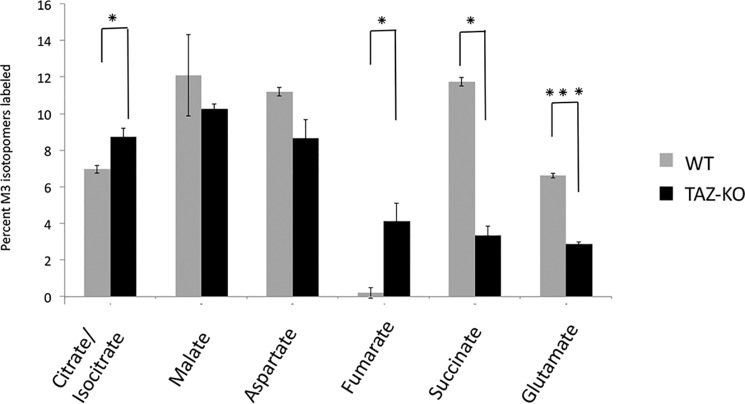
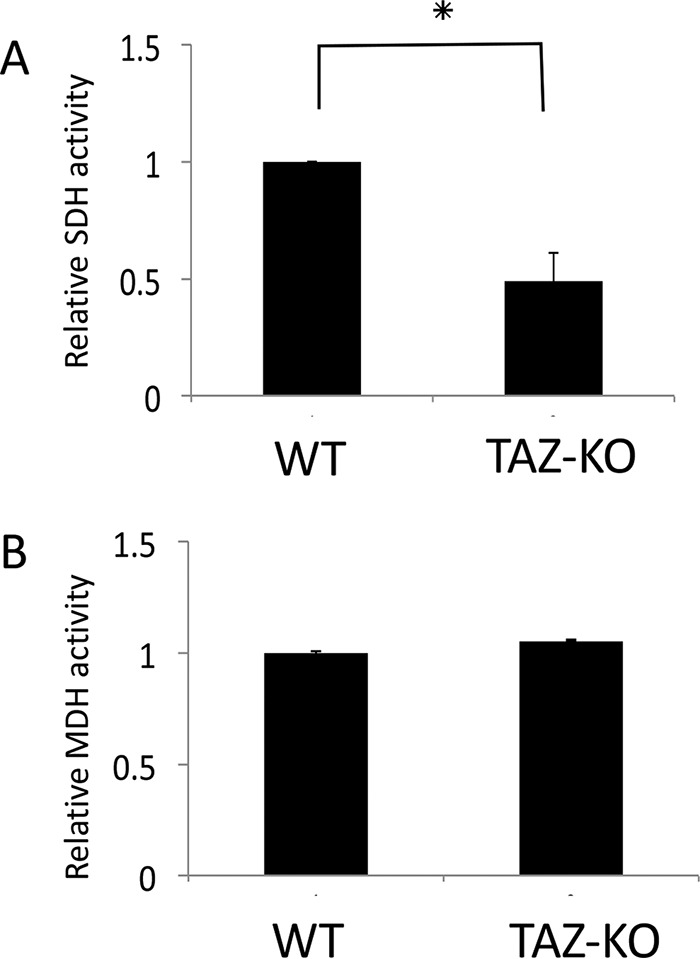
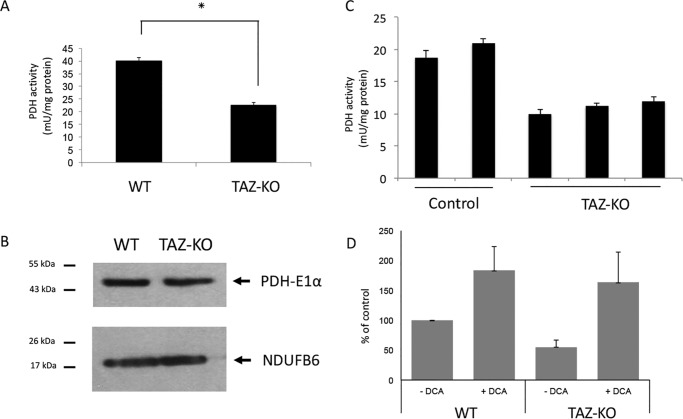
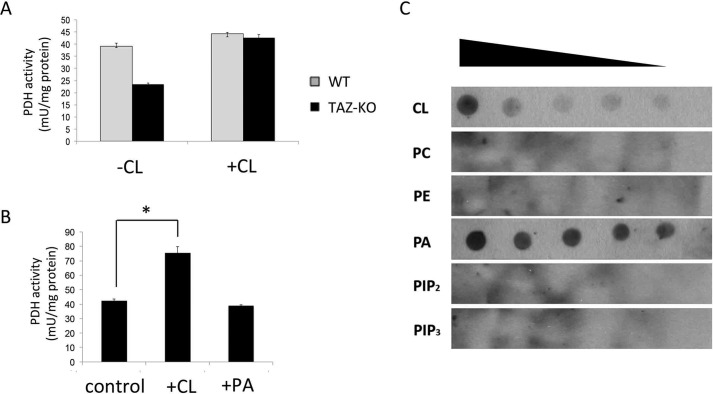
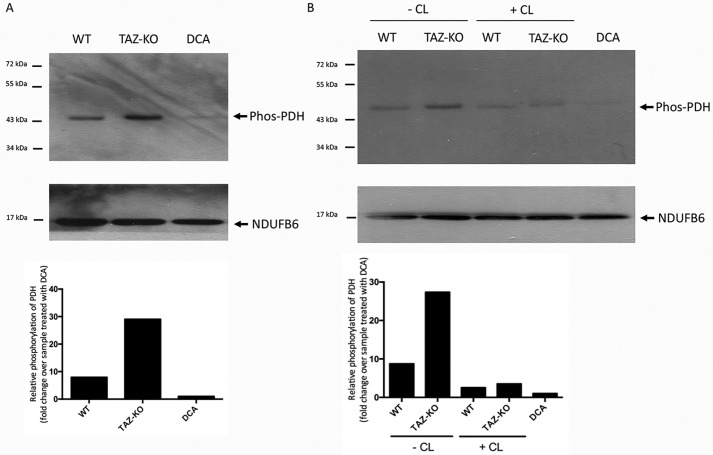
Cardiolipin (CL) is the signature phospholipid of mitochondrial membranes. Although it has long been known that CL plays an important role in mitochondrial bioenergetics, recent evidence in the yeast model indicates that CL is also essential for intermediary metabolism. To gain insight into the function of CL in energy metabolism in mammalian cells, here we analyzed the metabolic flux of [U-13C]glucose in a mouse C2C12 myoblast cell line, TAZ-KO, which is CL-deficient because of CRISPR/Cas9-mediated knockout of the CL-remodeling enzyme tafazzin (TAZ). TAZ-KO cells exhibited decreased flux of [U-13C]glucose to [13C]acetyl-CoA and M2 and M4 isotopomers of tricarboxylic acid (TCA) cycle intermediates. The activity of pyruvate carboxylase, the predominant enzyme for anaplerotic replenishing of the TCA cycle, was elevated in TAZ-KO cells, which also exhibited increased sensitivity to the pyruvate carboxylase inhibitor phenylacetate. We attributed a decreased carbon flux from glucose to acetyl-CoA in the TAZ-KO cells to a ∼50% decrease in pyruvate dehydrogenase (PDH) activity, which was observed in both TAZ-KO cells and cardiac tissue from TAZ-KO mice. Protein-lipid overlay experiments revealed that PDH binds to CL, and supplementing digitonin-solubilized TAZ-KO mitochondria with CL restored PDH activity to WT levels. Mitochondria from TAZ-KO cells exhibited an increase in phosphorylated PDH, levels of which were reduced in the presence of supplemented CL. These findings indicate that CL is required for optimal PDH activation, generation of acetyl-CoA, and TCA cycle function, findings that link the key mitochondrial lipid CL to TCA cycle function and energy metabolism.
Keywords: cardiolipin; mitochondria; pyruvate carboxylase (PC); pyruvate dehydrogenase complex (PDC); tricarboxylic acid cycle (TCA cycle) (Krebs cycle).
© 2019 Li et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
