

Rapid Evolution of Gained Essential Developmental Functions of a Young Gene via Interactions with Other Essential Genes
- PMID: 31187122
- PMCID: PMC6759199
- DOI: 10.1093/molbev/msz137
Rapid Evolution of Gained Essential Developmental Functions of a Young Gene via Interactions with Other Essential Genes
Abstract
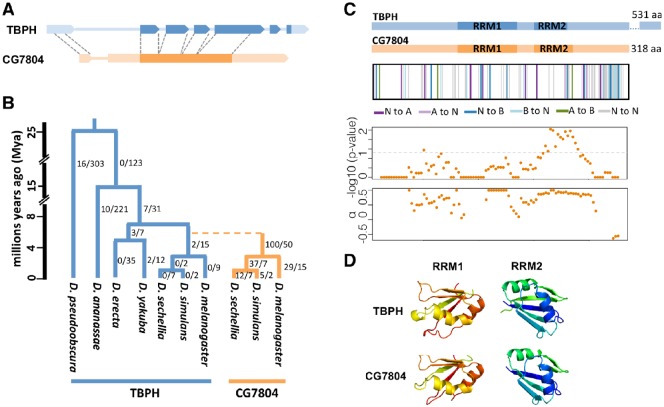
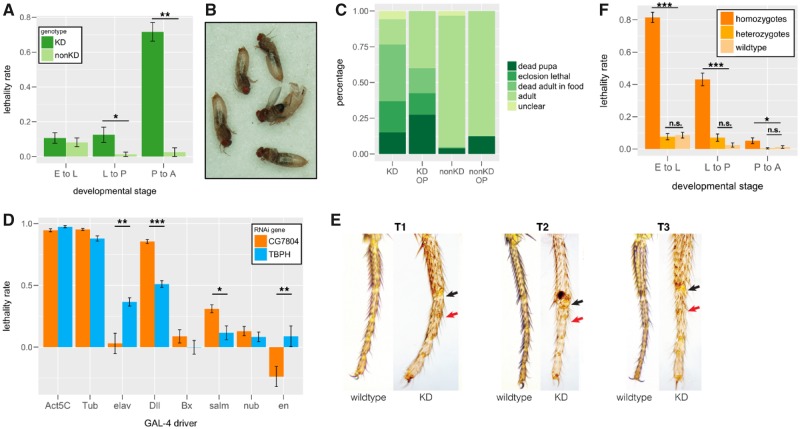
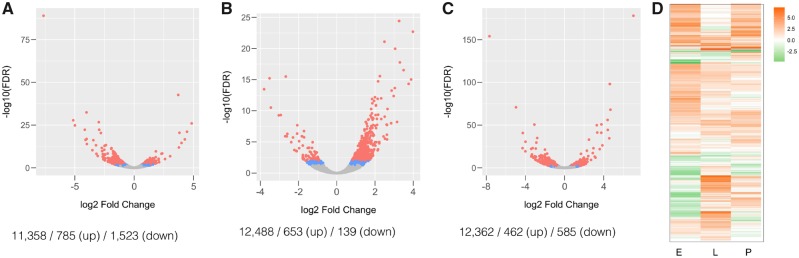
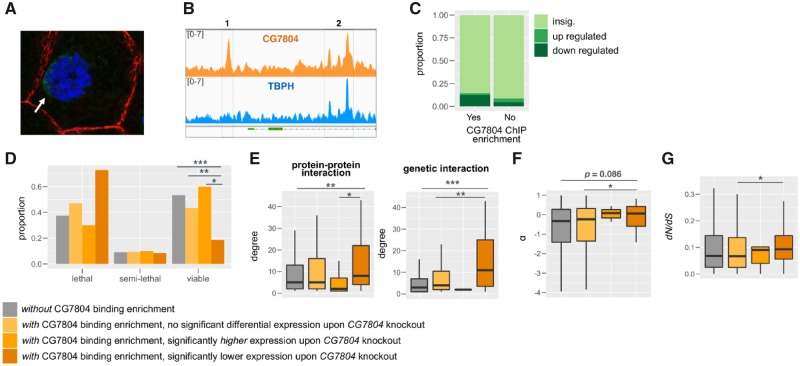
New genes are of recent origin and only present in a subset of species in a phylogeny. Accumulated evidence suggests that new genes, like old genes that are conserved across species, can also take on important functions and be essential for the survival and reproductive success of organisms. Although there are detailed analyses of the mechanisms underlying new genes' gaining fertility functions, how new genes rapidly become essential for viability remains unclear. We focused on a young retro-duplicated gene (CG7804, which we named Cocoon) in Drosophila that originated between 4 and 10 Ma. We found that, unlike its evolutionarily conserved parental gene, Cocoon has evolved under positive selection and accumulated many amino acid differences at functional sites from the parental gene. Despite its young age, Cocoon is essential for the survival of Drosophila melanogaster at multiple developmental stages, including the critical embryonic stage, and its expression is essential in different tissues from those of its parental gene. Functional genomic analyses found that Cocoon acquired unique DNA-binding sites and has a contrasting effect on gene expression to that of its parental gene. Importantly, Cocoon binding predominantly locates at genes that have other essential functions and/or have multiple gene-gene interactions, suggesting that Cocoon acquired novel essential function to survival through forming interactions that have large impacts on the gene interaction network. Our study is an important step toward deciphering the evolutionary trajectory by which new genes functionally diverge from parental genes and become essential.
Keywords: Drosophila; development; gene network; lethality; retrogene evolution.
© The Author(s) 2019. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
References
-
- Anon. Forthcoming. Babraham bioinformatics—trim galore! Available from: http://www.bioinformatics.babraham.ac.uk/projects/trim_galore/; Accessed March 2015.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
